

Regulation of TATA-binding protein binding by the SAGA complex and the Nhp6 high-mobility group protein
- PMID: 12612066
- PMCID: PMC149471
- DOI: 10.1128/MCB.23.6.1910-1921.2003
Regulation of TATA-binding protein binding by the SAGA complex and the Nhp6 high-mobility group protein
Abstract
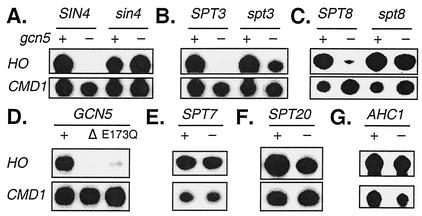
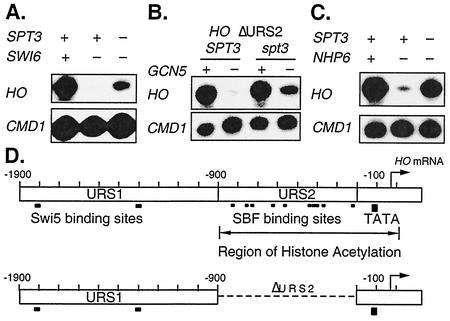
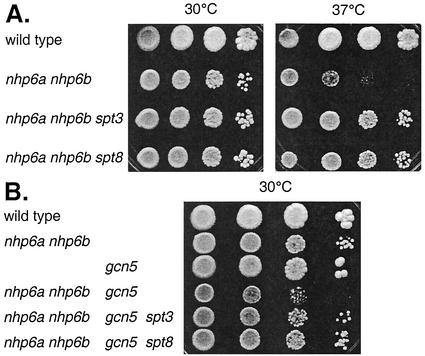
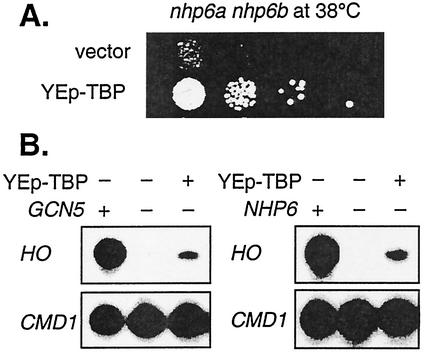
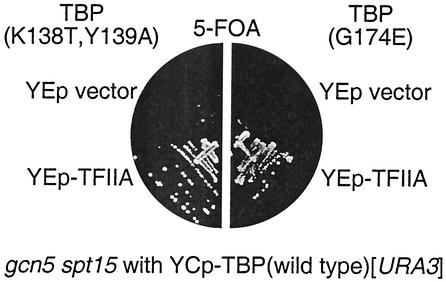
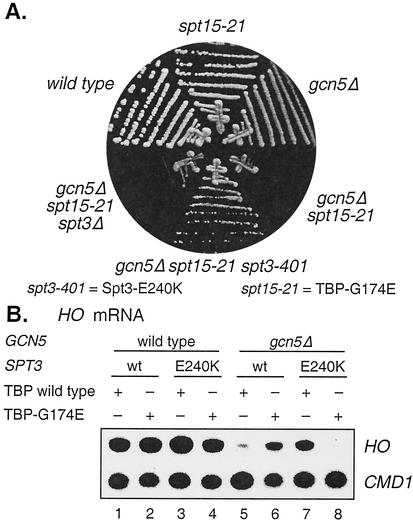
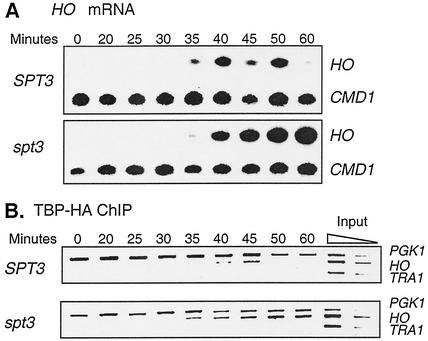
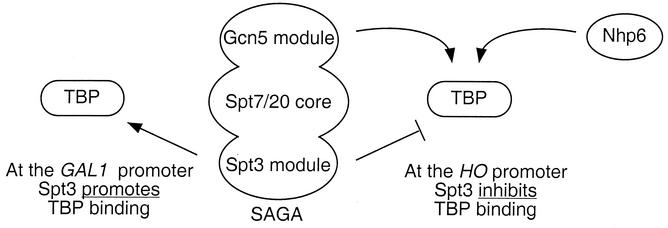
Transcriptional activation of the yeast HO gene involves the sequential action of DNA-binding and chromatin-modifying factors. Here we examine the role of the SAGA complex and the Nhp6 architectural transcription factor in HO regulation. Our data suggest that these factors regulate binding of the TATA-binding protein (TBP) to the promoter. A gcn5 mutation, eliminating the histone acetyltransferase present in SAGA, reduces the transcription of HO, but expression is restored in a gcn5 spt3 double mutant. We conclude that the major role of Gcn5 in HO activation is to overcome repression by Spt3. Spt3 is also part of SAGA, and thus two proteins in the same regulatory complex can have opposing roles in transcriptional regulation. Chromatin immunoprecipitation experiments show that TBP binding to HO is very weak in wild-type cells but markedly increased in an spt3 mutant, indicating that Spt3 reduces HO expression by inhibiting TBP binding. In contrast, it has been shown previously that Spt3 stimulates TBP binding to the GAL1 promoter as well as GAL1 expression, and thus, Spt3 regulates these promoters differently. We also find genetic interactions between TBP and either Gcn5 or the high-mobility-group protein Nhp6, including multicopy suppression and synthetic lethality. These results suggest that, while Spt3 acts to inhibit TBP interaction with the HO promoter, Gcn5 and Nhp6 act to promote TBP binding. The result of these interactions is to limit TBP binding and HO expression to a short period within the cell cycle. Furthermore, the synthetic lethality resulting from combining a gcn5 mutation with specific TBP point mutations can be suppressed by the overexpression of transcription factor IIA (TFIIA), suggesting that histone acetylation by Gcn5 can stimulate transcription by promoting the formation of a TBP/TFIIA complex.
Figures

Similar articles
-
SWI/SNF binding to the HO promoter requires histone acetylation and stimulates TATA-binding protein recruitment.Mol Cell Biol. 2006 Jun;26(11):4095-110. doi: 10.1128/MCB.01849-05. Mol Cell Biol. 2006. PMID: 16705163 Free PMC article.
-
Role for Nhp6, Gcn5, and the Swi/Snf complex in stimulating formation of the TATA-binding protein-TFIIA-DNA complex.Mol Cell Biol. 2004 Sep;24(18):8312-21. doi: 10.1128/MCB.24.18.8312-8321.2004. Mol Cell Biol. 2004. PMID: 15340090 Free PMC article.
-
Genetic interactions between Nhp6 and Gcn5 with Mot1 and the Ccr4-Not complex that regulate binding of TATA-binding protein in Saccharomyces cerevisiae.Genetics. 2006 Feb;172(2):837-49. doi: 10.1534/genetics.105.050245. Epub 2005 Nov 4. Genetics. 2006. PMID: 16272410 Free PMC article.
-
Architecture of the multi-functional SAGA complex and the molecular mechanism of holding TBP.FEBS J. 2021 May;288(10):3135-3147. doi: 10.1111/febs.15563. Epub 2020 Sep 29. FEBS J. 2021. PMID: 32946670 Review.
-
Role of the TATA-box binding protein (TBP) and associated family members in transcription regulation.Gene. 2022 Jul 30;833:146581. doi: 10.1016/j.gene.2022.146581. Epub 2022 May 18. Gene. 2022. PMID: 35597524 Review.
Cited by
-
Requirement of Nhp6 proteins for transcription of a subset of tRNA genes and heterochromatin barrier function in Saccharomyces cerevisiae.Mol Cell Biol. 2007 Mar;27(5):1545-57. doi: 10.1128/MCB.00773-06. Epub 2006 Dec 18. Mol Cell Biol. 2007. PMID: 17178828 Free PMC article.
-
Facilitated assembly of the preinitiation complex by separated tail and head/middle modules of the mediator.J Mol Biol. 2012 Jan 20;415(3):464-74. doi: 10.1016/j.jmb.2011.11.020. Epub 2011 Nov 23. J Mol Biol. 2012. PMID: 22137896 Free PMC article.
-
SWI/SNF binding to the HO promoter requires histone acetylation and stimulates TATA-binding protein recruitment.Mol Cell Biol. 2006 Jun;26(11):4095-110. doi: 10.1128/MCB.01849-05. Mol Cell Biol. 2006. PMID: 16705163 Free PMC article.
-
High throughput screening identifies modulators of histone deacetylase inhibitors.BMC Genomics. 2014 Jun 26;15(1):528. doi: 10.1186/1471-2164-15-528. BMC Genomics. 2014. PMID: 24968945 Free PMC article.
-
Snf1p regulates Gcn5p transcriptional activity by antagonizing Spt3p.Genetics. 2010 Jan;184(1):91-105. doi: 10.1534/genetics.109.110957. Epub 2009 Oct 19. Genetics. 2010. PMID: 19841091 Free PMC article.
References
-
- Agalioti, T., S. Lomvardas, B. Parekh, J. Yie, T. Maniatis, and D. Thanos. 2000. Ordered recruitment of chromatin modifying and general transcription factors to the IFN-beta promoter. Cell 103:667-678. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials