

Induction of primary virus-cross-reactive human immunodeficiency virus type 1-neutralizing antibodies in small animals by using an alphavirus-derived in vivo expression system
- PMID: 12584337
- PMCID: PMC149731
- DOI: 10.1128/jvi.77.5.3119-3130.2003
Induction of primary virus-cross-reactive human immunodeficiency virus type 1-neutralizing antibodies in small animals by using an alphavirus-derived in vivo expression system
Abstract
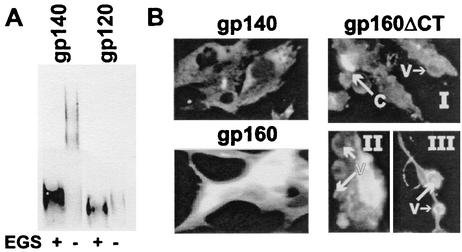
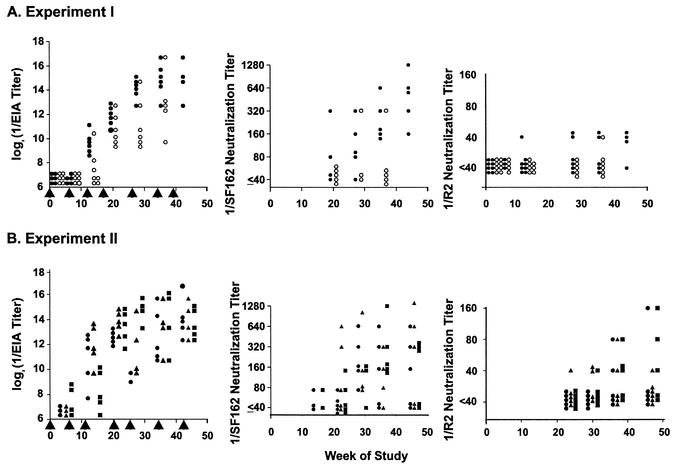
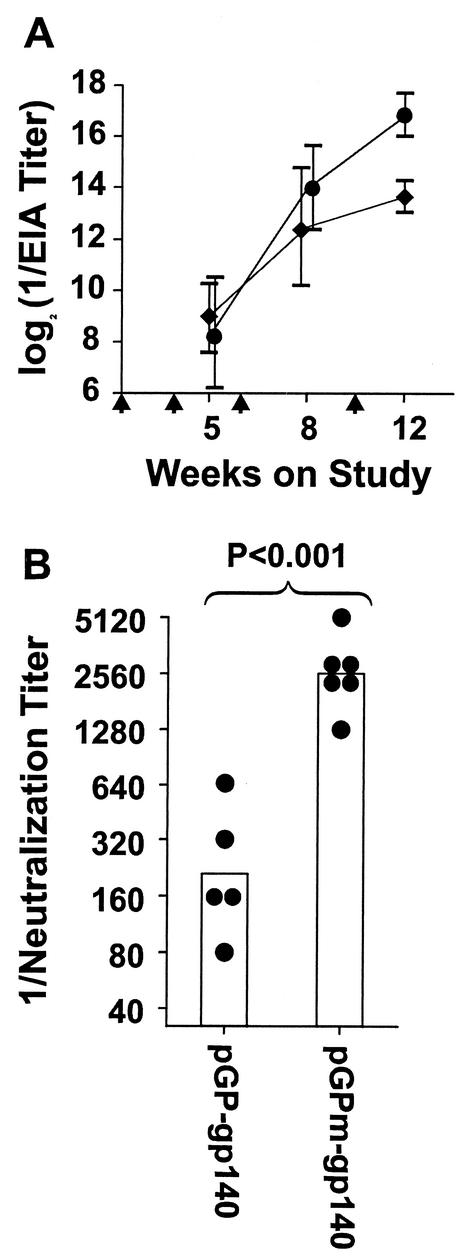
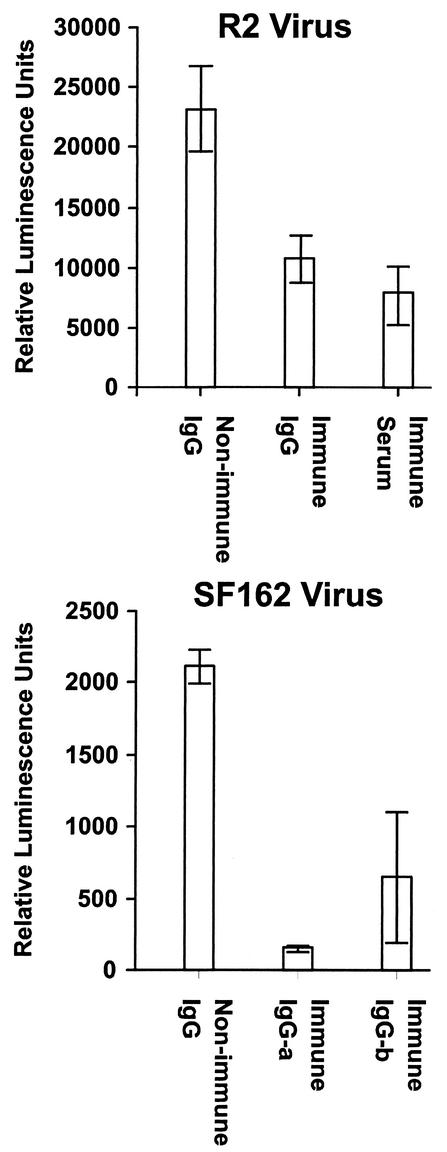
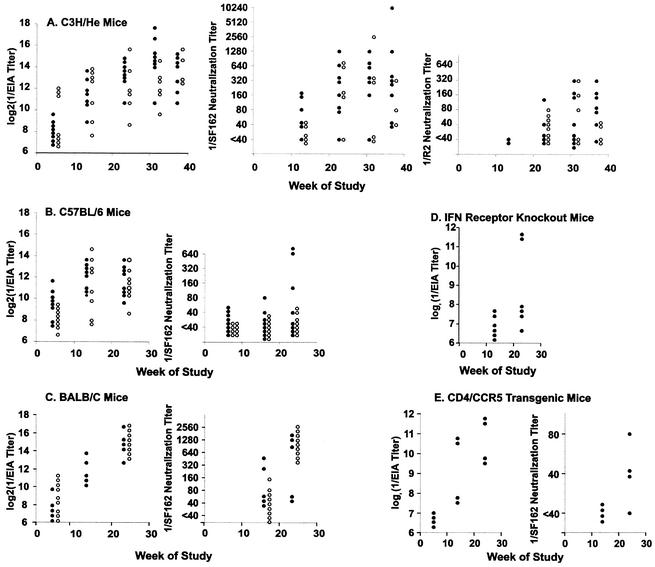
We have studied the induction of neutralizing antibodies by in vivo expression of the human immunodeficiency virus type 1 (HIV-1) envelope by using a Venezuelan equine encephalitis virus (VEE) replicon system with mice and rabbits. The HIV-1 envelope, clone R2, has broad sensitivity to cross-reactive neutralization and was obtained from a donor with broadly cross-reactive, primary virus-neutralizing antibodies (donor of reference serum, HIV-1-neutralizing serum 2 [HNS2]). It was expressed as gp160, as secreted gp140, and as gp160deltaCT with the cytoplasmic tail deleted. gp140 was expressed in vitro at a high level and was predominantly uncleaved oligomer. gp160deltaCT was released by cells in the form of membrane-bound vesicles. gp160deltaCT induced stronger neutralizing responses than the other forms. Use of a helper plasmid for replicon particle packaging, in which the VEE envelope gene comprised a wild-type rather than a host range-adapted sequence, also enhanced immunogenicity. Neutralizing activity fractionated with immunoglobulin G. This activity was cross-reactive among a panel of five nonhomologous primary clade B strains and a Chinese clade C strain and minimally reactive against a Chinese clade E (circulating recombinant form 1) strain. The comparative neutralization of these strains by immune mouse sera was similar to the relative neutralizing effects of HNS2, and responses induced in rabbits were similar to those induced in mice. Together, these results demonstrate that neutralizing antibody responses can be induced in mice within 2 to 3 months that are similar in potency and cross-reactivity to those found in the chronically infected, long-term nonprogressive donor of HNS2. These findings support the expectation that induction of highly cross-reactive HIV-1 primary virus-neutralizing activity by vaccination may be realized.
Figures
Similar articles
-
Evaluation of envelope vaccines derived from the South African subtype C human immunodeficiency virus type 1 TV1 strain.J Virol. 2005 Nov;79(21):13338-49. doi: 10.1128/JVI.79.21.13338-13349.2005. J Virol. 2005. PMID: 16227256 Free PMC article.
-
A variable region 3 (V3) mutation determines a global neutralization phenotype and CD4-independent infectivity of a human immunodeficiency virus type 1 envelope associated with a broadly cross-reactive, primary virus-neutralizing antibody response.J Virol. 2002 Jan;76(2):644-55. doi: 10.1128/jvi.76.2.644-655.2002. J Virol. 2002. PMID: 11752155 Free PMC article.
-
Protection of rhesus monkeys against infection with minimally pathogenic simian-human immunodeficiency virus: correlations with neutralizing antibodies and cytotoxic T cells.J Virol. 2005 Mar;79(6):3358-69. doi: 10.1128/JVI.79.6.3358-3369.2005. J Virol. 2005. PMID: 15731230 Free PMC article.
-
Alphavirus replicon particles as candidate HIV vaccines.IUBMB Life. 2002 Apr-May;53(4-5):209-11. doi: 10.1080/15216540212657. IUBMB Life. 2002. PMID: 12120997 Review.
-
Relevance of the antibody response against human immunodeficiency virus type 1 envelope to vaccine design.Immunol Lett. 1997 Jun 1;57(1-3):105-12. doi: 10.1016/s0165-2478(97)00043-6. Immunol Lett. 1997. Corrected and republished in: Immunol Lett. 1997 Jul;58(2):125-32. doi: 10.1016/s0165-2478(97)00109-0 PMID: 9232434 Corrected and republished. Review.
Cited by
-
A group M consensus envelope glycoprotein induces antibodies that neutralize subsets of subtype B and C HIV-1 primary viruses.Virology. 2006 Sep 30;353(2):268-82. doi: 10.1016/j.virol.2006.04.043. Virology. 2006. PMID: 17039602 Free PMC article.
-
Immunogenicity of virus-like particles containing modified human immunodeficiency virus envelope proteins.Vaccine. 2007 May 10;25(19):3841-50. doi: 10.1016/j.vaccine.2007.01.107. Epub 2007 Feb 15. Vaccine. 2007. PMID: 17320250 Free PMC article.
-
Soluble HIV-1 envelope immunogens derived from an elite neutralizer elicit cross-reactive V1V2 antibodies and low potency neutralizing antibodies.PLoS One. 2014 Jan 23;9(1):e86905. doi: 10.1371/journal.pone.0086905. eCollection 2014. PLoS One. 2014. PMID: 24466285 Free PMC article.
-
Lack of interference with immunogenicity of a chimeric alphavirus replicon particle-based influenza vaccine by preexisting antivector immunity.Clin Vaccine Immunol. 2012 Jul;19(7):991-8. doi: 10.1128/CVI.00031-12. Epub 2012 May 23. Clin Vaccine Immunol. 2012. PMID: 22623651 Free PMC article.
-
Evolution of antibody landscape and viral envelope escape in an HIV-1 CRF02_AG infected patient with 4E10-like antibodies.Retrovirology. 2009 Dec 14;6:113. doi: 10.1186/1742-4690-6-113. Retrovirology. 2009. PMID: 20003438 Free PMC article.
References
-
- Beirnaert, E., P. Nyambi, B. Willems, L. Heyndrickx, R. Colebunders, W. Janssens, and G. van Der. 2000. Identification and characterization of sera from HIV-infected individuals with broad cross-neutralizing activity against group M (env clade A-H) and group O primary HIV-1 isolates. J. Med. Virol. 62:14-24. - PubMed
-
- Caley, I. J., M. R. Betts, N. L. Davis, R. Swanstrom, J. A. Frelinger, and R. E. Johnston. 1999. Venezuelan equine encephalitis virus vectors expressing HIV-1 proteins: vector design strategies for improved vaccine efficacy. Vaccine 17:3124-3135. - PubMed
-
- Caley, I. J., M. R. Betts, D. M. Irlbeck, N. L. Davis, R. Swanstrom, J. A. Frelinger, and R. E. Johnston. 1997. Humoral, mucosal, and cellular immunity in response to a human immunodeficiency virus type 1 immunogen expressed by a Venezuelan equine encephalitis virus vaccine vector. J. Virol. 71:3031-3038. - PMC - PubMed
-
- Charles, P. C., K. W. Brown, N. L. Davis, M. K. Hart, and R. E. Johnston. 1997. Mucosal immunity induced by parenteral immunization with a live attenuated Venezuelan equine encephalitis virus vaccine candidate. Virology 228:153-160. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
