

Enhanced muscle fat oxidation and glucose transport by ACRP30 globular domain: acetyl-CoA carboxylase inhibition and AMP-activated protein kinase activation
- PMID: 12456889
- PMCID: PMC138607
- DOI: 10.1073/pnas.222657499
Enhanced muscle fat oxidation and glucose transport by ACRP30 globular domain: acetyl-CoA carboxylase inhibition and AMP-activated protein kinase activation
Abstract
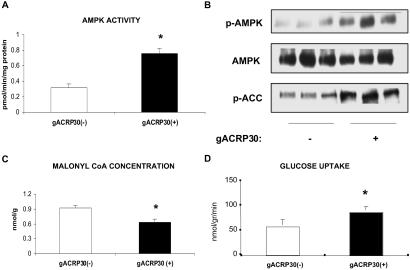
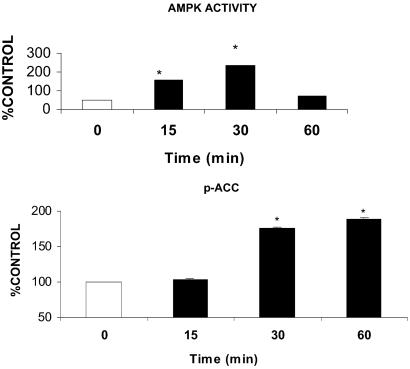
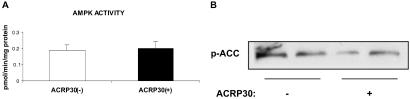
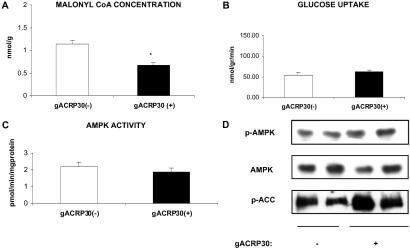
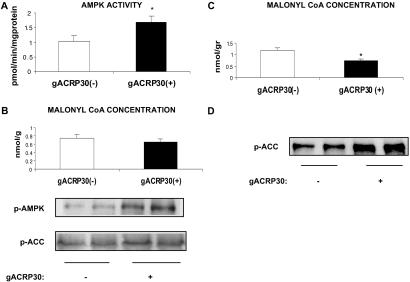
gACRP30, the globular subunit of adipocyte complement-related protein of 30 kDa (ACRP30), improves insulin sensitivity and increases fatty acid oxidation. The mechanism by which gACRP30 exerts these effects is unknown. Here, we examined if gACRP30 activates AMP-activated protein kinase (AMPK), an enzyme that has been shown to increase muscle fatty acid oxidation and insulin sensitivity. Incubation of rat extensor digitorum longus (EDL), a predominantly fast twitch muscle, with gACRP30 (2.5 micro g/ml) for 30 min led to 2-fold increases in AMPK activity and phosphorylation of both AMPK on Thr-172 and acetyl CoA carboxylase (ACC) on Ser-79. Accordingly, concentration of malonyl CoA was diminished by 30%. In addition, gACRP30 caused a 1.5-fold increase in 2-deoxyglucose uptake. Similar changes in malonyl CoA and ACC were observed in soleus muscle incubated with gACRP30 (2.5 micro g/ml), although no significant changes in AMPK activity or 2-deoxyglucose uptake were detected. When EDL was incubated with full-length hexameric ACRP30 (10 micro g/ml), AMPK activity and ACC phosphorylation were not altered. Administration of gACRP30 (75 micro g) to C57 BL6J mice in vivo led to increased AMPK activity and ACC phosphorylation and decreased malonyl CoA concentration in gastrocnemius muscle within 15-30 min. Both in vivo and in vitro, activation of AMPK was the first effect of gACRP30 and was transient, whereas alterations in malonyl CoA and ACC occurred later and were more sustained. Thus, gACRP30 most likely exerts its actions on muscle fatty acid oxidation by inactivating ACC via activation of AMPK and perhaps other signal transduction proteins.
Figures
Similar articles
-
LKB1 and the regulation of malonyl-CoA and fatty acid oxidation in muscle.Am J Physiol Endocrinol Metab. 2007 Dec;293(6):E1572-9. doi: 10.1152/ajpendo.00371.2007. Epub 2007 Oct 9. Am J Physiol Endocrinol Metab. 2007. PMID: 17925454
-
Effect of phosphorylation by AMP-activated protein kinase on palmitoyl-CoA inhibition of skeletal muscle acetyl-CoA carboxylase.J Appl Physiol (1985). 2005 Apr;98(4):1221-7. doi: 10.1152/japplphysiol.00621.2004. Epub 2004 Dec 3. J Appl Physiol (1985). 2005. PMID: 15579580
-
AMPK and ACC phosphorylation: effect of leptin, muscle fibre type and obesity.Mol Cell Endocrinol. 2008 Mar 12;284(1-2):1-10. doi: 10.1016/j.mce.2007.12.013. Epub 2008 Jan 3. Mol Cell Endocrinol. 2008. PMID: 18255222
-
AMP-activated protein kinase regulation of fatty acid oxidation in the ischaemic heart.Biochem Soc Trans. 2003 Feb;31(Pt 1):207-12. doi: 10.1042/bst0310207. Biochem Soc Trans. 2003. PMID: 12546686 Review.
-
Malonyl CoA control of fatty acid oxidation in the ischemic heart.J Mol Cell Cardiol. 2002 Sep;34(9):1099-109. doi: 10.1006/jmcc.2002.2060. J Mol Cell Cardiol. 2002. PMID: 12392882 Review.
Cited by
-
Serum Adiponectin Levels as an Independent Marker of Severity of Psoriasis: A Cross-Sectional Analysis.J Psoriasis Psoriatic Arthritis. 2023 Oct;8(4):148-155. doi: 10.1177/24755303231199995. Epub 2023 Aug 31. J Psoriasis Psoriatic Arthritis. 2023. PMID: 39301471 Free PMC article.
-
Physiological Appetite Regulation and Bariatric Surgery.J Clin Med. 2024 Feb 27;13(5):1347. doi: 10.3390/jcm13051347. J Clin Med. 2024. PMID: 38546831 Free PMC article. Review.
-
Decreased AdipoR1 signaling and its implications for obesity-induced male infertility.Sci Rep. 2024 Mar 8;14(1):5701. doi: 10.1038/s41598-024-56290-0. Sci Rep. 2024. PMID: 38459078 Free PMC article.
-
Antidiabetic features of AdipoAI, a novel AdipoR agonist.Cell Biochem Funct. 2024 Jan;42(1):e3910. doi: 10.1002/cbf.3910. Cell Biochem Funct. 2024. PMID: 38269524
-
Goldfish adiponectin: (I) molecular cloning, tissue distribution, recombinant protein expression, and novel function as a satiety factor in fish model.Front Endocrinol (Lausanne). 2023 Oct 30;14:1283298. doi: 10.3389/fendo.2023.1283298. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 38027109 Free PMC article.
References
-
- Tsao T. S., Lodish, H. F. & Fruebis, J. (2002) Eur. J. Pharmacol. 440, 213-221. - PubMed
-
- Scherer P. E., Williams, S., Fogliano, M., Baldini, G. & Lodish, H. F. (1995) J. Biol. Chem. 270, 26746-26749. - PubMed
-
- Tsao T. S., Murrey, H. E., Hug, C., Lee, D. H. & Lodish, H. F. (2002) J. Biol. Chem. 277, 29359-29362. - PubMed
-
- Arita Y., Kihara, S., Ouchi, N., Takahashi, M., Meada, K., Miyagawa, J., Hotta, K., Shimomura, I., Nakamura, I., Miyaoka, K., et al. (1999) Biochem. Biophys. Res. Commun. 257, 79-83. - PubMed
-
- Hotta K., Funahashi, T., Arita, Y., Takahashi, M., Matsuda, M., Okamoto, Y., Iwahashi, H., Kuriyama, H., Ouchi, N., Maeda, K., et al. (2000) Arterioscler. Thromb. Vasc. Biol. 20, 1595-1599. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
