

doi: 10.1083/jcb.200208154.
Epub 2002 Nov 18.
Membrane proximal lysosomes are the major vesicles responsible for calcium-dependent exocytosis in nonsecretory cells
Affiliations
- PMID: 12438417
- PMCID: PMC2173094
- DOI: 10.1083/jcb.200208154
Item in Clipboard
Membrane proximal lysosomes are the major vesicles responsible for calcium-dependent exocytosis in nonsecretory cells
J Cell Biol.
.
Abstract
Similar to its role in secretory cells, calcium triggers exocytosis in nonsecretory cells. This calcium-dependent exocytosis is essential for repair of membrane ruptures. Using total internal reflection fluorescence microscopy, we observed that many organelles implicated in this process, including ER, post-Golgi vesicles, late endosomes, early endosomes, and lysosomes, were within 100 nm of the plasma membrane (in the evanescent field). However, an increase in cytosolic calcium led to exocytosis of only the lysosomes. The lysosomes that fused were predominantly predocked at the plasma membrane, indicating that calcium is primarily responsible for fusion and not recruitment of lysosomes to the cell surface.
Figures

Distribution of cellular organelles in the evanescent field. CHO cells were transiently transfected with (A) Calreticulin-EYFP; (B) hGH-EGFP; (C) GalT-ECFP; (D) VAMP8-ECFP; (E) Rab7-ECFP; and (F) CD63-EGFP and imaged using epifluorescence (left) and TIR-FM (right). TIR-FM visualizes the structures that are within 100 nm of the plasma membrane. Note that GalT-ECFP staining (C) shows a total absence of the Golgi apparatus near the plasma membrane. Bar, 10 μm.

Lysosomes are located within 100 nm of the plasma membrane in several cell types. (A) Long-term loading with dextran specifically labels the lysosome. Endocytic compartments of CHO cells transiently transfected with VAMP-ECFP, Rab7-ECFP, SytVII-EGFP, VAMP7-ECFP, and CD63-EGFP were loaded for 2 h with TRITC dextran or Texas red dextran followed by a 5-h chase in dextran-free media. The images represent 0.4 μm thick Z confocal sections which contained the maximum number of labeled vesicles. (B) Lysosomes are detected by TIR close to the plasma membrane. CHO cells transiently transfected with SytVII-EGFP, Lamp1-EGFP, and VAMP7-ECFP were imaged using TIR-FM. Each of these lysosome specific markers were found to be present within the evanescent field. (C) Lysosomes are detected by TIR close to the plasma membrane in several cell types. Lysosomes in NIH3T3 fribroblasts, WM239 melanoma cells, HeLa cells, MDCK epithelial cells, and murine embryonic primary fibroblasts were loaded with FITC dextran as in Fig. 1, and the cells were visualized using TIR-FM. Bar, 10 μm.

Lysosomes are located within 100 nm of the plasma membrane in several cell types. (A) Long-term loading with dextran specifically labels the lysosome. Endocytic compartments of CHO cells transiently transfected with VAMP-ECFP, Rab7-ECFP, SytVII-EGFP, VAMP7-ECFP, and CD63-EGFP were loaded for 2 h with TRITC dextran or Texas red dextran followed by a 5-h chase in dextran-free media. The images represent 0.4 μm thick Z confocal sections which contained the maximum number of labeled vesicles. (B) Lysosomes are detected by TIR close to the plasma membrane. CHO cells transiently transfected with SytVII-EGFP, Lamp1-EGFP, and VAMP7-ECFP were imaged using TIR-FM. Each of these lysosome specific markers were found to be present within the evanescent field. (C) Lysosomes are detected by TIR close to the plasma membrane in several cell types. Lysosomes in NIH3T3 fribroblasts, WM239 melanoma cells, HeLa cells, MDCK epithelial cells, and murine embryonic primary fibroblasts were loaded with FITC dextran as in Fig. 1, and the cells were visualized using TIR-FM. Bar, 10 μm.

Lysosomal membrane and lumenal markers confirm that calcium induces exocytosis of lysosomes. NRK cells labeled with either CD63-EGFP (A–C) or CD63-EGFP (D) and 10 kD Texas red–dextran were treated with 10 μm A23187 ionophore and imaged using TIR-FM. A region from A that shows a cell with CD63-EGFP–labeled lysosomes in the evanescent field. A lysosome that underwent exocytosis has been enlarged in B; the arrow indicates a CD63-EGFP–labeled lysosome that moved into the evanescent field during the period of observation and fused to the plasma membrane. Note that it takes ∼3 s for the fluorescence to diffuse out along the membrane. (C) The quantification of the peak, total intensity and (half-width)2 of the fluorescence for a lysosome that undergoes fusion (the values for peak intensity has been multiplied by a factor of five). The point when lysosome ceased moving before initiation of fusion was used as t = 0. (D) Representative images of a lysosome double labeled for membrane (green; using CD63-EGFP) and lumen (red; Texas red dextran) that underwent calcium-induced fusion. The arrow points to a lysosome that underwent exocytosis. The time stamp represents the time elapsed after the addition of ionophore. Note that the dextran fluorescene (red) diffuses much faster compared with the diffusion of the fluorescence of the membrane marker (green; Video 2).

Characterization of lysosomal exocytosis by monitoring the release of a lumenal marker. Lysosomes in NRK cells were loaded with FITC dextran and cells were treated with 10 μm A23187 calcium-ionophore. Although the epifluorescence image (A) predominantly demonstrates the perinuclear population of lysosomes, TIR-FM (B) allows identification of lysosomes near the plasma membrane. In response to the addition of calcium ionophore there is a reduction in lysosomal movement and ∼7% of the total lysosomes in the evanescent field exocytose (Video 3). (C) Most of the exocytosing lysosomes are predocked (arrowhead), whereas a few recent arrivals (arrow) also exocytose. Time stamp indicates the time elapsed since the addition of ionophore. (D) Quantification of changes in the total intensity and (half-width)2 of the fluorescence of exocytosing dextran. X-axis represents the time starting from when the lysosome ceased moving before initiation of fusion. Note that the spread phase for dextran (a lumenal marker) is much shorter compared with the spread phase for a CD63-EGFP (a membrane marker; Fig. 3 C).
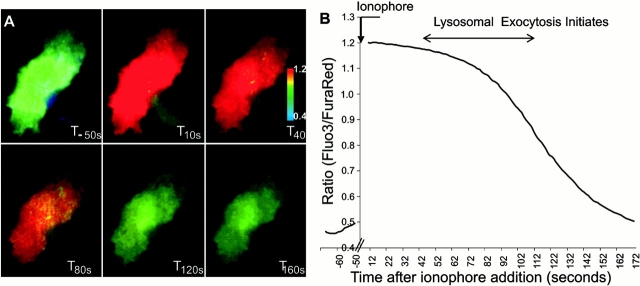
Lysosomal exocytosis initiates with a delay after the increase in intracellular calcium. CHO cells were loaded for 20 min with Fluo-3-AM and Fura red–AM, after which 10μm A23187 calcium-ionophore was added (T0s) and the cells were imaged using TIR-FM. The cells were illuminated using 488 nm laser light and the emission from both the two dyes was simultaneously and continuously imaged as described in Materials and methods. The images were subtracted for background and pseudo colored ratio images of Fluo-3/Fura red were generated. (A) Selected ratio images of a cell at various time points after the addition of ionophore. Within 10 s after the addition of ionophore, calcium reached the peak value, which dropped to prestimulatory levels over next 120 s. The pseudo color code represents increasing ratio (hence calcium) from blue to red (B) The trace of ratio of Fluo-3/Fura red emission intensities before and after the addition of ionophore. Note that in all the cell lines imaged (n = 42 cells), lysosomal exocytosis was initiated during the phase when submembrane calcium level started to decline, after the initial ionophore induced increase.

Predocked lysosomes are primarily responsible for calcium induced exocytosis. Cells loaded with fluorescent dextran were treated with 10 μM calcium-ionophore, 0.2 U/ml thrombin, or 20 nM bombesin. The cells were continuously imaged using TIR-FM. A frame-by-frame analysis was performed to identify the exocytosing and motile lysosomes. In the case of both (A) CHO cells and (B) murine embryonic primary fibroblasts, treatment with calcium ionophore, thrombin and bombesin led to lysosomal exocytosis. The murine embryonic primary fibroblasts showed approximately fivefold greater exocytosis than the CHO cells. (C) Increase in calcium caused both by ionophore and IP3 agonists, bombesin and thrombin led to a reduced motility of the lysosomes in the evanescent field. (Lysosomes that appeared or disappeared from the evanescent field, or moved >1 μm within the evanescent field were classified as motile.)
Similar articles
-
Differential properties of GTP- and Ca(2+)-stimulated exocytosis from large dense core vesicles.Traffic. 2006 Apr;7(4):416-28. doi: 10.1111/j.1600-0854.2006.00394.x. Traffic. 2006. PMID: 16536740
-
Visualization of regulated exocytosis with a granule-membrane probe using total internal reflection microscopy.Mol Biol Cell. 2004 Oct;15(10):4658-68. doi: 10.1091/mbc.e04-02-0149. Epub 2004 Jul 28. Mol Biol Cell. 2004. PMID: 15282339 Free PMC article.
-
Lysosomes behave as Ca2+-regulated exocytic vesicles in fibroblasts and epithelial cells.J Cell Biol. 1997 Apr 7;137(1):93-104. doi: 10.1083/jcb.137.1.93. J Cell Biol. 1997. PMID: 9105039 Free PMC article.
-
Lysosomes and plasma membrane repair.Curr Top Membr. 2019;84:1-16. doi: 10.1016/bs.ctm.2019.08.001. Epub 2019 Sep 3. Curr Top Membr. 2019. PMID: 31610859 Review.
-
There's more to life than neurotransmission: the regulation of exocytosis by synaptotagmin VII.Trends Cell Biol. 2005 Nov;15(11):626-31. doi: 10.1016/j.tcb.2005.09.001. Epub 2005 Sep 15. Trends Cell Biol. 2005. PMID: 16168654 Review.
Cited by
-
Evidence for lysosomal exocytosis and release of aggrecan-degrading hydrolases from hypertrophic chondrocytes, in vitro and in vivo.Biol Open. 2012 Apr 15;1(4):318-28. doi: 10.1242/bio.2012547. Epub 2012 Feb 10. Biol Open. 2012. PMID: 23213422 Free PMC article.
-
Rab27a and Rab27b regulate neutrophil azurophilic granule exocytosis and NADPH oxidase activity by independent mechanisms.Traffic. 2010 Apr;11(4):533-47. doi: 10.1111/j.1600-0854.2009.01029.x. Epub 2009 Dec 17. Traffic. 2010. PMID: 20028487 Free PMC article.
-
Palmitoylation-dependent association with CD63 targets the Ca2+ sensor synaptotagmin VII to lysosomes.J Cell Biol. 2010 Nov 1;191(3):599-613. doi: 10.1083/jcb.201003021. J Cell Biol. 2010. PMID: 21041449 Free PMC article.
-
The secreted inhibitor of invasive cell growth CREG1 is negatively regulated by cathepsin proteases.Cell Mol Life Sci. 2021 Jan;78(2):733-755. doi: 10.1007/s00018-020-03528-5. Epub 2020 May 8. Cell Mol Life Sci. 2021. PMID: 32385587 Free PMC article.
-
Role of AP1 and Gadkin in the traffic of secretory endo-lysosomes.Mol Biol Cell. 2011 Jun 15;22(12):2068-82. doi: 10.1091/mbc.E11-03-0193. Epub 2011 Apr 27. Mol Biol Cell. 2011. PMID: 21525240 Free PMC article.
References
-
- Ayala, B.P., B. Vasquez, S. Clary, J.A. Tainer, K. Rodland, and M. So. 2001. The pilus-induced Ca2+ flux triggers lysosome exocytosis and increases the amount of Lamp1 accessible to Neisseria IgA1 protease. Cell Microbiol. 3:265–275. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
