

Review
doi: 10.1038/nrm934.
Furin at the cutting edge: from protein traffic to embryogenesis and disease
Affiliations
- PMID: 12360192
- PMCID: PMC1964754
- DOI: 10.1038/nrm934
Item in Clipboard
Review
Furin at the cutting edge: from protein traffic to embryogenesis and disease
Nat Rev Mol Cell Biol.
2002 Oct.
Abstract
Furin catalyses a simple biochemical reaction--the proteolytic maturation of proprotein substrates in the secretory pathway. But the simplicity of this reaction belies furin's broad and important roles in homeostasis, as well as in diseases ranging from Alzheimer's disease and cancer to anthrax and Ebola fever. This review summarizes various features of furin--its structural and enzymatic properties, intracellular localization, trafficking, substrates, and roles in vivo.
Figures
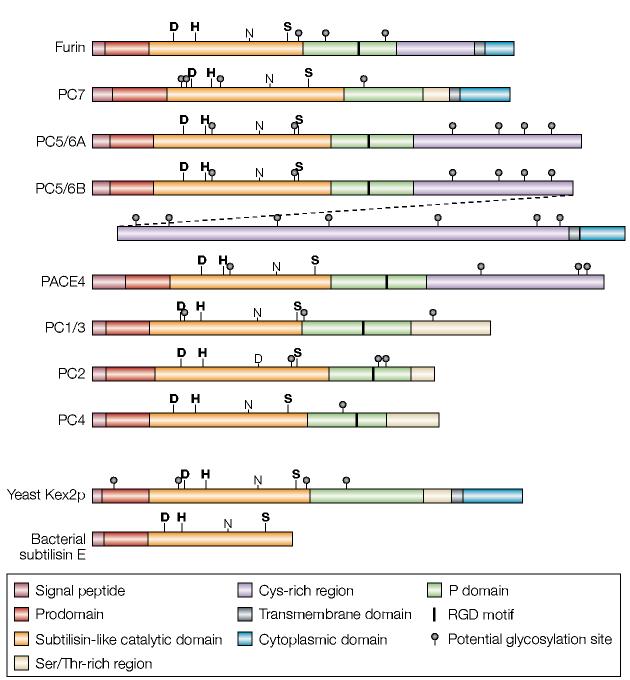
Schematic diagram of the proprotein convertase (PC) family. Shown are schematics for furin and the six other PCs. Schematics of yeast Kex2 and the evolutionarily related bacterial subtilisin (which lacks the conserved P domain) are also shown. PC5/6 is expressed as either the A or B isoform. These isoforms are generated by alternative splicing, and the diagonal dashed line links the two halves of PC5/6B. The PC5/6B isoform contains all of PC5/6A except for a small part of its carboxyl terminus that is positioned after the splice site. The bold labels D, H and S highlight the active-site residues, whereas the non-bold labels N and D highlight the oxyanion-hole residues.
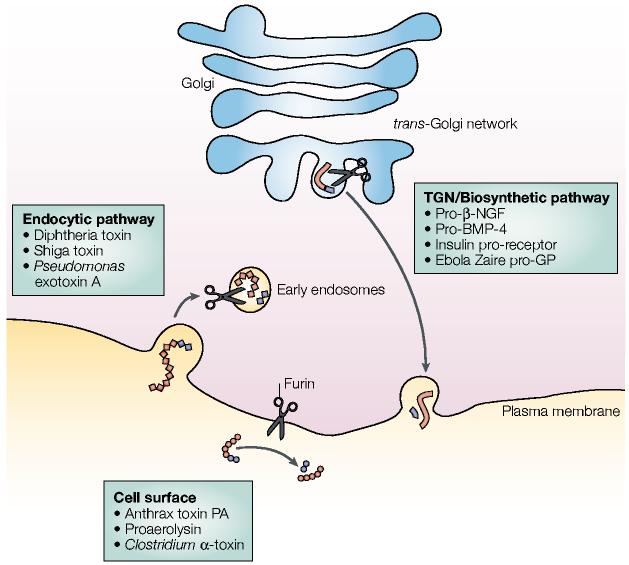
Furin-processing compartments of the trans-Golgi network (TGN)/endosomal system. At steady-state, furin (represented by scissors) is localized principally to the TGN, where it cycles between this sorting compartment, the cell surface and the early endosomes. In the TGN/biosynthetic pathway, furin cleaves many substrates including pro-β-nerve growth factor (pro-β-NGF), pro-bone morphogenetic protein-4 (pro-BMP-4), the insulin pro-receptor and Ebola Zaire pro-glycoprotein (pro-GP). At the cell surface, furin cleaves substrates such as anthrax protective antigen (PA), proaerolysin and Clostridium septicum α-toxin. In mildly acidic early endosomes (endocytic pathway), furin cleaves substrates including diphtheria toxins, shiga toxin and shiga-like toxin-1, and Pseudomonas exotoxin A. See text for more details.
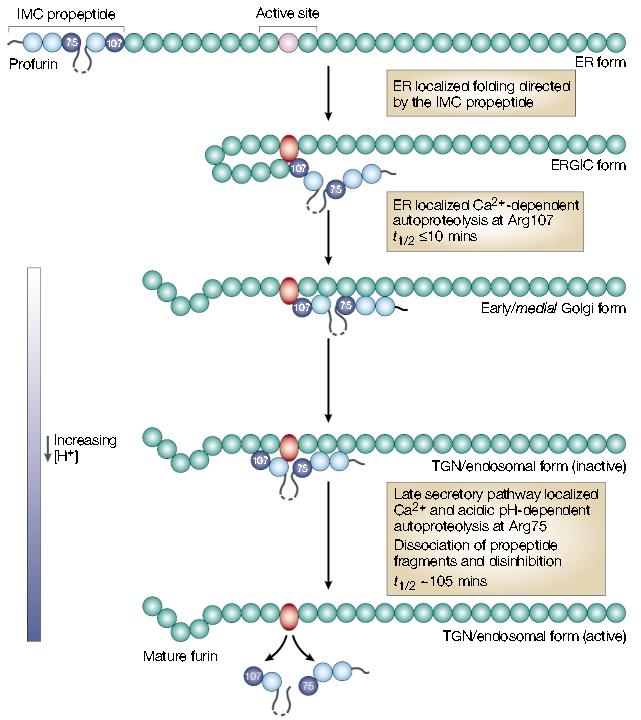
The furin autoactivation pathway. Following translocation and signal sequence removal, the furin prodomain acts as an intramolecular chaperone (IMC) to facilitate folding of the unstructured, inactive catalytic domain (pink circle) into the active conformation (red oval). After the initial endoplasmic reticulum (ER) folding events, furin undergoes autoproteolytic intramolecular excision of the propeptide at Arg107. The propeptide, however, remains associated with the mature domain and functions as a potent autoinhibitor in trans during transport to the late secretory pathway. Propeptide excision can be blocked by inactivating furin and results in the accumulation of an apparent folding intermediate in the ER–Golgi intermediate compartment (ERGIC)/cis-Golgi network. These data indicate that both the ER and ERGIC compartments participate in the initial steps of furin activation. Following propeptide excision, the inactive propeptide complex transits to late secretory compartments — trans-Golgi network (TGN)/endosomes — where the relatively acidic pH promotes autoproteolytic, intramolecular cleavage of the propeptide at a second, internal site (Arg75). The Arg75 cleavage is followed by the rapid dissociation of the propeptide fragments and disinhibition of furin. Adapted with permission from ref . . © (2002) American Society for Biochemistry and Molecular Biology.
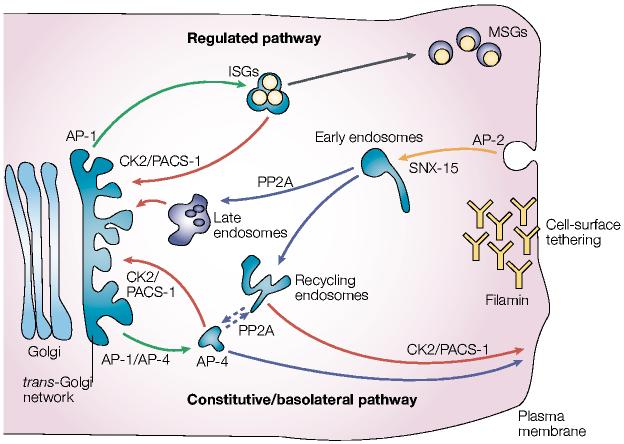
Model of furin trafficking. Budding of furin from the trans-Golgi network (TGN) is mediated by the binding of the tyrosine-based or di-leucine-like hydrophobic sorting motifs to adaptor protein (AP)-1, which targets furin to endosomes, or binding to AP-4, which targets furin to the basolateral surface from either the TGN or possibly endosomes. In endocrine and neuroendocrine cells, AP-1 directs furin budding from the TGN into immature secretory granules (ISGs). Phosphofurin acidic cluster sorting protein-1 (PACS-1) connects the casein kinase 2 (CK2)-phosphorylated furin acidic cluster to AP-1/clathrin to retrieve furin to the TGN either from ISGs, prior to their progression to mature secretory granules (MSGs), or from endosomes. Furin molecules arriving at the cell surface can be tethered by the cytoskeletal protein filamin, which is also called actin-binding protein (ABP)-280. The dynamin/clathrin-dependent internalization of cell-surface furin is mediated principally by the tyrosine-based motif, which binds to AP-2. Once inside early endosomes, furin molecules that are dephosphorylated by specific protein phosphatase 2A (PP2A) isoforms are delivered to the TGN, apparently through a late endosomal compartment. By contrast, CK2-phosphorylated furin is recycled back to the plasma membrane in a PACS-1-dependent step. Hence, the TGN- and peripheral-cycling loops are essentially mirror images of each other. On the basis of the similar itinery of carboxypeptidase D (CPD), movement of furin from a post-TGN endosomal compartment to the cell surface might also require PP2A (dashed lines). Sorting nexin-15 (SNX-15), a PX-domain containing protein that binds phosphoinositides, modulates furin sorting through endosomes.
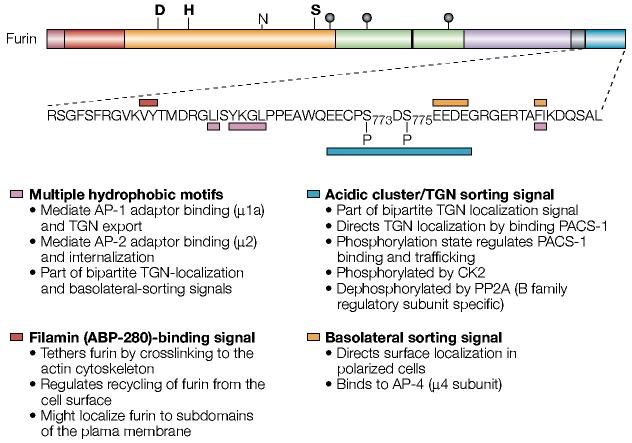
The sorting motifs of the furin cytoplasmic domain. Shown is the sequence of the human furin cytoplasmic domain. The various intracellular sorting motifs are indicated. See text for more details. ABP, actin-binding protein; AP, adaptor protein; CK2, casein kinase 2; PACS-1, phosphofurin acidic cluster sorting protein-1; PP2A, protein phosphatase 2A; TGN, trans-Golgi network. For a key to furin domain organization, see FIG. 1.
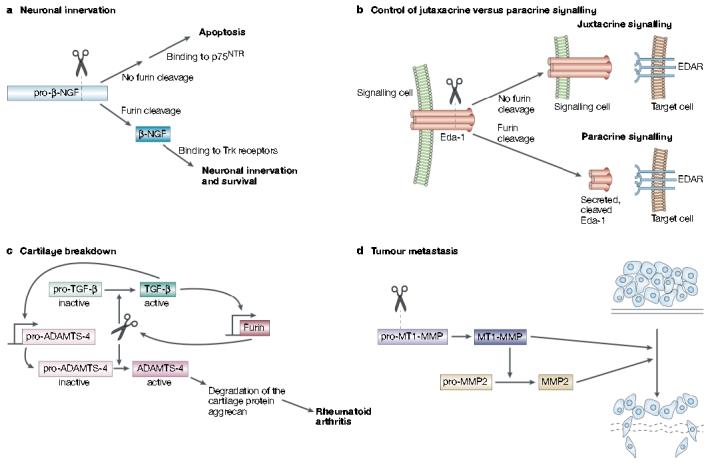
Furin in development, homeostasis and disease. a | Furin-mediated cleavage of pro-β-nerve growth factor (NGF) produces the 13-kDa β-NGF neurotrophin that binds to Trk receptors to promote synaptic innervation. By contrast, inhibition or sequestering of furin results in the secretion of pro-β-NGF that binds to the 75-kDa neurotrophin receptor (p75NTR) to promote cell-death pathways. b | Ectodysplasin-A (Eda-1) is a trimeric tumour necrosis factor family member that stimulates morphogenesis of ectodermal structures by activation of its receptor, EDAR, on target cells. Eda-1 can signal in a juxtacrine manner by binding to EDAR on adjacent cells. However, cleavage of membrane anchored Eda-1 by furin releases the ligand and enables it to signal through EDAR on distant cells in a paracrine manner. c | In synoviocytes, furin and transforming growth factor (TGF)-β participate in a positive feedback loop that results in elevated levels of ‘a disintegrin and metalloprotease with thrombospondin motifs-4’ (ADAMTS-4, or aggrecanase-1). Furin cleaves both pro-TGF-β and pro-ADAMTS-4 to yield the active growth factor and protease, respectively. The secreted mature form of TGF-β then binds to its receptor and, through a SMAD2 and mitogen-activated protein kinase (MAPK) convergent pathway, increases furin expression. The increased levels of furin lead to an increase in TGF-β, which creates a positive-feedback loop. In synoviocytes, TGF-β also stimulates the expression of pro-ADAMTS-4. So, because furin and TGF-β are in this positive loop, the levels of active ADAMTS-4 are greatly elevated, which leads to destruction of the cartilage protein aggrecan and hence to rheumatoid arthritis. d | Furin activates several membrane-type matrix metalloproteinases (MT-MMPs) that are involved in tumour formation and metastasis. Furin-activated MT-MMP1 activates MMP2 (gelatinase), which degrades the extracellular matrix, and MT-MMP1 also directly degrades extracellular matrix itself.
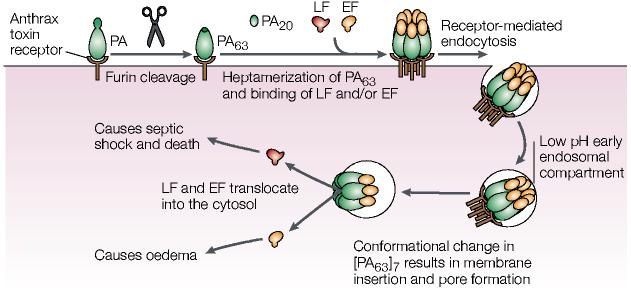
Furin activation of the anthrax toxin. Cleavage of anthrax protective antigen (PA) by furin leads to internalization and activation of lethal factor (LF), which is a zinc metalloproteinase that cleaves mitogen-activated protein kinase (MAPK) kinases, and oedema factor (EF), which is a calmodulin-dependent adenylate cyclase. EF causes eschar formation, which are black masses of scab-like necrotized tissue, and massive soft-tissue oedema. In lung macrophages, phagocytosed LF leads to changes in the levels of tumour necrosis factor-α and interleukin-1β, as well as apoptosis. The altered cytokine levels coupled with the paralysed innate immune system rapidly result in anthrax-induced systemic shock and death by a complex, unresolved process. See text for more details. Modified with permission from REF. . © (2002) Birkhäuser Publishing Ltd.
Similar articles
-
Bi-cycling the furin pathway: from TGN localization to pathogen activation and embryogenesis.Trends Cell Biol. 1999 Jan;9(1):28-35. doi: 10.1016/s0962-8924(98)01382-8. Trends Cell Biol. 1999. PMID: 10087614 Review.
-
Cytoskeletal protein ABP-280 directs the intracellular trafficking of furin and modulates proprotein processing in the endocytic pathway.J Cell Biol. 1997 Dec 29;139(7):1719-33. doi: 10.1083/jcb.139.7.1719. J Cell Biol. 1997. PMID: 9412467 Free PMC article.
-
The crystal structure of the proprotein processing proteinase furin explains its stringent specificity.Nat Struct Biol. 2003 Jul;10(7):520-6. doi: 10.1038/nsb941. Nat Struct Biol. 2003. PMID: 12794637
-
Activation of membrane-type matrix metalloproteinase 3 zymogen by the proprotein convertase furin in the trans-Golgi network.Cancer Res. 2002 Feb 1;62(3):675-81. Cancer Res. 2002. PMID: 11830519
-
Structure and function of eukaryotic proprotein processing enzymes of the subtilisin family of serine proteases.Crit Rev Oncog. 1993;4(2):115-36. Crit Rev Oncog. 1993. PMID: 8420571 Review.
Cited by
-
Regulation of ADAM10 activity through microdomain-dependent intracellular calcium changes.Cell Commun Signal. 2024 Nov 4;22(1):531. doi: 10.1186/s12964-024-01891-5. Cell Commun Signal. 2024. PMID: 39497138 Free PMC article.
-
Cytomegalovirus Generates Assembly Compartment in the Early Phase of Infection by Perturbation of Host-Cell Factors Recruitment at the Early Endosome/Endosomal Recycling Compartment/Trans-Golgi Interface.Front Cell Dev Biol. 2020 Sep 11;8:563607. doi: 10.3389/fcell.2020.563607. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 33042998 Free PMC article.
-
Assembly and Cellular Exit of Coronaviruses: Hijacking an Unconventional Secretory Pathway from the Pre-Golgi Intermediate Compartment via the Golgi Ribbon to the Extracellular Space.Cells. 2021 Feb 26;10(3):503. doi: 10.3390/cells10030503. Cells. 2021. PMID: 33652973 Free PMC article. Review.
-
SARS-CoV-2 cellular tropism and direct multiorgan failure in COVID-19 patients: Bioinformatic predictions, experimental observations, and open questions.Cell Biol Int. 2023 Feb;47(2):308-326. doi: 10.1002/cbin.11928. Epub 2022 Oct 13. Cell Biol Int. 2023. PMID: 36229927 Free PMC article. Review.
-
Protection of the Furin Cleavage Site in Low-Toxicity Immunotoxins Based on Pseudomonas Exotoxin A.Toxins (Basel). 2016 Jul 25;8(8):217. doi: 10.3390/toxins8080217. Toxins (Basel). 2016. PMID: 27463727 Free PMC article.
References
-
- Seidah NG, Day R, Marcinkiewicz M, Chretien M. Precursor convertases: an evolutionary ancient, cell-specific, combinatorial mechanism yielding diverse bioactive peptides and proteins. Ann. N. Y. Acad. Sci. 1998;839:9–24. - PubMed
-
- Thacker C, Rose AM. A look at the Caenorhabditis elegans Kex2/Subtilisin-like proprotein convertase family. Bioessays. 2000;22:545–553. - PubMed
-
- Zhou A, Martin S, Lipkind G, LaMendola J, Steiner DF. Regulatory roles of the P domain of the subtilisin-like prohormone convertases. J. Biol. Chem. 1998;273:11107–11114. - PubMed
-
- Siezen RJ, Creemers JW, Van de Ven WJ. Homology modelling of the catalytic domain of human furin. A model for the eukaryotic subtilisin-like proprotein convertases. Eur. J. Biochem. 1994;222:255–266. - PubMed
-
- Rockwell NC, Fuller RS. Specific modulation of Kex2/Furin family proteases by potassium. J. Biol. Chem. 2002;277:17531–17537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
