

doi: 10.1073/pnas.202200399.
Epub 2002 Sep 23.
An activation domain in the C-terminal subunit of HCF-1 is important for transactivation by VP16 and LZIP
Affiliations
- PMID: 12271126
- PMCID: PMC129685
- DOI: 10.1073/pnas.202200399
Item in Clipboard
An activation domain in the C-terminal subunit of HCF-1 is important for transactivation by VP16 and LZIP
Proc Natl Acad Sci U S A.
.
Abstract
In herpes simplex virus, lytic replication is initiated by the viral transactivator VP16 acting with cellular cofactors Oct-1 and HCF-1. Although this activator complex has been studied in detail, the role of HCF-1 remains elusive. Here, we show that HCF-1 contains an activation domain (HCF-1(AD)) required for maximal transactivation by VP16 and its cellular counterpart LZIP. Expression of the VP16 cofactor p300 augments HCF-1(AD) activity, suggesting a mechanism of synergy. Infection of cells lacking the HCF-1(AD) leads to reduced viral immediate-early gene expression and lowered viral titers. These findings underscore the importance of HCF-1 to herpes simplex virus replication and VP16 transactivation.
Figures
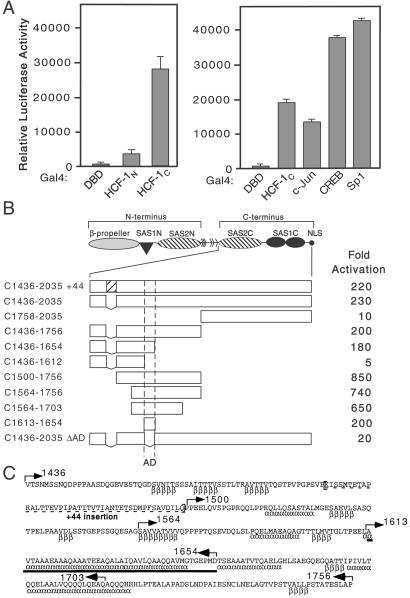
The C terminus of HCF-1 contains an activation domain. (A) 293T cells were transfected with expression plasmids (250 ng) encoding Gal4DBD, Gal4-HCF-1N, Gal4-HCF-1C, or the activation domains of Gal4-c-Jun, Gal4-CREB, and Gal4-Sp1, and assayed for transactivation of a 5xGal4-E1B-luc reporter (500 ng). Extracts were prepared after 40 h and assayed for luciferase activity. Values represent means and standard deviations of three independent transfections. (B) The structure of the HCF-1 precursor is shown. The boundaries of the activation domain were delineated using N-terminal, C-terminal, or internal deletions fused to the Gal4DBD. Luciferase activity is scored as fold activation above Gal4DBD alone. (C) Sequence of human HCF-1 (residues 1436–1756) encompassing the HCF-1AD (solid bar). Alternative splicing removes 44 amino acids (dashed underline). Secondary structure predictions [α (α-helix) and β (β-sheet)] were derived using the Frishman and Argos method (50). Arrows indicate the boundaries of truncations used in B.
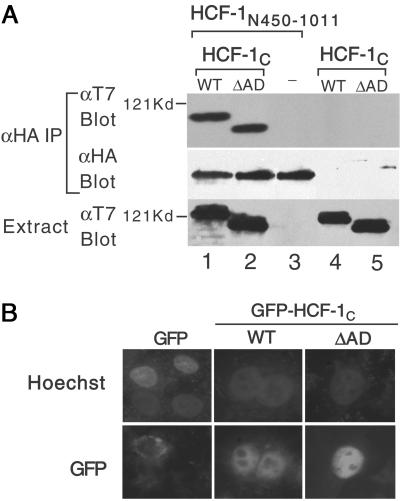
Deletion of HCF-1AD does not affect association with the N terminus or nuclear localization. (A) HA-tagged HCF-1N450–1011 and T7-tagged HCF-1C WT or ΔAD were coexpressed in transfected 293T cells and association assayed by immunoprecipitation with αHA antibody followed by immunoblotting with the αT7 antibody. The HA-tagged HCF-1N450–1011 is in lanes 1–3 and the T7-tagged HCF-1C WT and ΔAD in lanes 1 and 4 and lanes 2 and 5, respectively. (B) HeLa cells were transfected with expression plasmids (500 ng) encoding GFP or GFP-tagged HCF-1C WT or ΔAD. DNA is visualized with Hoechst 33258 and GFP by fluorescence microscopy.
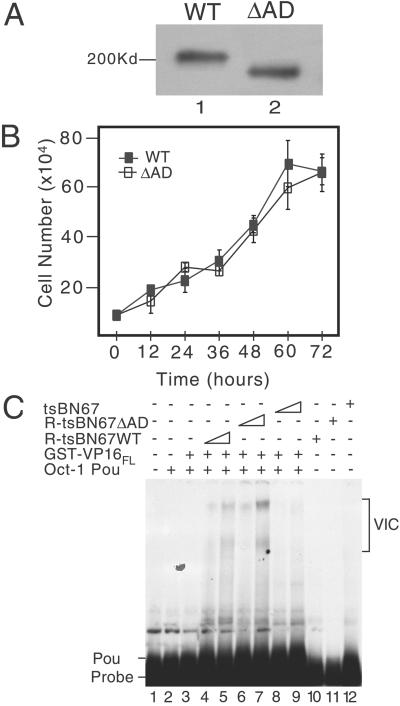
Characterization of R-tsBN67 HCF-1Δrep WT and ΔAD cells. (A) Immunoblot comparing tsBN67-derived cell lines stably expressing T7-tagged HCF-1Δrep WT and ΔAD. Protein extracts were prepared from equal numbers of R-tsBN67-HCF-1Δrep WT (lane 1) and ΔAD (lane 2) cells, resolved on a SDS-7% polyacrylamide gel and immunoblotted with the αT7 antibody. (B) Analysis of cell proliferation. R-tsBN67-HCF-1Δrep WT (■) and ΔAD (□) cells were seeded at a density of 1 × 105 cells per 10-cm dish and incubated at 39.5°C for 3 days. At the indicated times, three separate dishes were harvested and counted. (C) Protein extracts were prepared from an equal number of tsBN67, R-tsBN67-HCF-1ΔrepWT, and HCF-1ΔrepΔAD cells and assayed for the ability to support VP16-induced complex formation in a gel-mobility shift assay using a 32P-labeled ICP0 TAATGARAT element as a probe. Lane 1 contains probe alone, lanes 2–9 contain probe with Oct-1 POU domain, and lanes 3–9 contain probe, Oct-1 POU domain, and VP16FL fused to GST. Two amounts of each cell extract (a 3-fold difference) were incubated with VP16 and Oct-1 POU: WT extract (lanes 4 and 5), ΔAD extract (lanes 6 and 7), and tsBN67 extract (lanes 8 and 9). In lanes 10, 11, and 12, the probe was incubated with the highest amount of WT, ΔAD, and tsBN67 extracts.
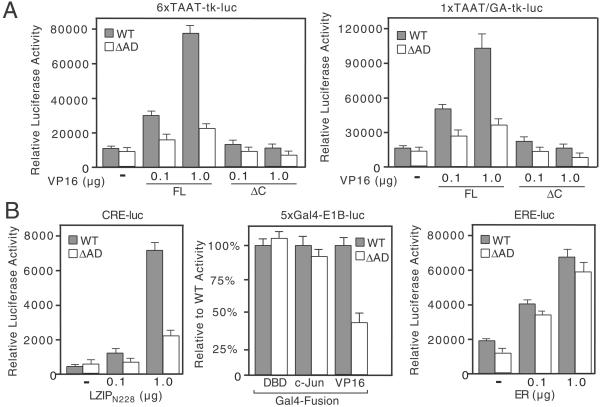
Transactivation by VP16 and LZIP is severely compromised in tsBN67-derived cells expressing HCF-1ΔrepΔAD. (A) R-tsBN67-HCF-1ΔrepWT and R-tsBN67-HCF-1ΔrepΔAD cells were transiently transfected with 6xTAAT-tk-luc (500 ng) or 1xTAAT/GA-tk-luc (1 μg) reporters together with 0.1 or 1.0 μg of the VP16FL or VP16ΔC expression plasmids and assayed for luciferase activity after incubation at 39.5°C for 40 h. (B) As in A, except cells were transfected with: (Left) a CRE-luciferase reporter (CRE-luc, 500 ng) and an expression plasmid encoding LZIPN228; (Center) a Gal4-responsive reporter (5xGal4-E1B-luc reporter, 500 ng) and 500 ng of either Gal4DBD, Gal4-c-Jun, or Gal4-VP16AD; or (Right) an estrogen response element reporter (ERE-luc, 500 ng) and plasmid encoding the human estrogen receptor (ER).
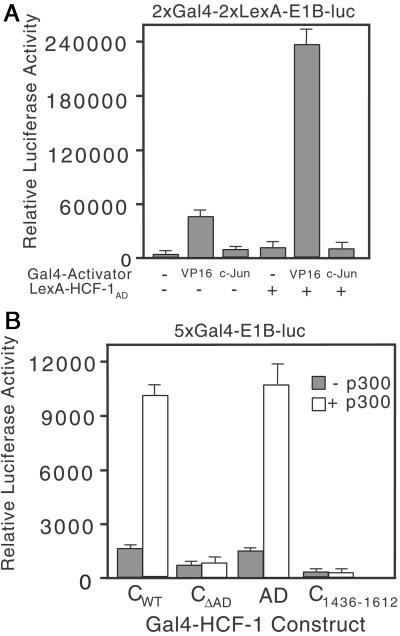
The HCF-1AD synergizes with VP16 and p300. (A) 293T cells were transfected with 2xGal4-2xLexA-E1B-luc (200 ng) together with Gal4 activator (VP16AD or c-Jun, 100 ng) and LexA-HCF-1AD (10 ng). Extracts were prepared after 40 h and assayed for luciferase activity. (B) 293T cells were cotransfected with expression plasmids (500 ng) encoding Gal4-HCF-1CWT or ΔAD, Gal4-HCF-1C1436–1612, or Gal4-HCF-1AD with the 5xGal4-E1B-luc reporter gene (500 ng) and with (open bars) or without (filled bars) an expression plasmid (6 μg) encoding full-length p300.
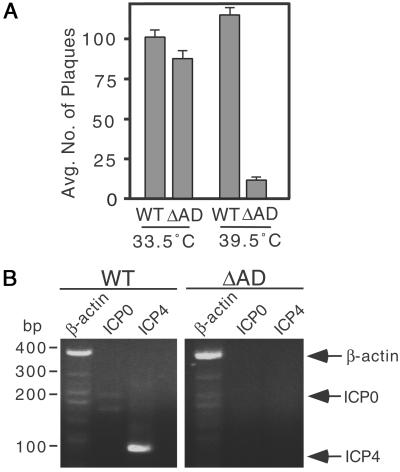
The HCF-1AD is involved in HSV lytic replication. (A) Wild-type HSV-1 at moi of 0.001 was used to infect R-tsBN67-HCF-1Δrep WT and ΔAD cells maintained at 33.5°C and 39.5°C. After 5 days, cells were harvested, sonicated, and serial dilutions plated on Vero cells. After 4 days, cells were fixed and stained with crystal violet to visualize viral plaques. Graph shows the mean number of plaques at 10−6 the dilution based on four independent assays. (B) Wild-type HSV-1 at moi of 0.001 was used to infect R-tsBN67-HCF-1Δrep WT and ΔAD cells maintained at 39.5°C. RNA was isolated after 4 h and assayed by RT-PCR. Specific amplification products from hamster β-actin (lanes 1 and 4, amplification fragment, 360 bp), ICP0 (lanes 2 and 5, amplification fragment, 157 bp), and ICP4 (lanes 3 and 6 amplification fragment, 101 bp) are indicated.
Similar articles
-
N-terminal transcriptional activation domain of LZIP comprises two LxxLL motifs and the host cell factor-1 binding motif.Proc Natl Acad Sci U S A. 2000 Sep 26;97(20):10757-62. doi: 10.1073/pnas.190062797. Proc Natl Acad Sci U S A. 2000. PMID: 10984507 Free PMC article.
-
Mutations in host cell factor 1 separate its role in cell proliferation from recruitment of VP16 and LZIP.Mol Cell Biol. 2000 Feb;20(3):919-28. doi: 10.1128/MCB.20.3.919-928.2000. Mol Cell Biol. 2000. PMID: 10629049 Free PMC article.
-
Zhangfei: a second cellular protein interacts with herpes simplex virus accessory factor HCF in a manner similar to Luman and VP16.Nucleic Acids Res. 2000 Jun 15;28(12):2446-54. doi: 10.1093/nar/28.12.2446. Nucleic Acids Res. 2000. PMID: 10871379 Free PMC article.
-
The herpes simplex virus VP16-induced complex: the makings of a regulatory switch.Trends Biochem Sci. 2003 Jun;28(6):294-304. doi: 10.1016/S0968-0004(03)00088-4. Trends Biochem Sci. 2003. PMID: 12826401 Review.
-
The herpes simplex virus VP16-induced complex: mechanisms of combinatorial transcriptional regulation.Cold Spring Harb Symp Quant Biol. 1998;63:599-607. doi: 10.1101/sqb.1998.63.599. Cold Spring Harb Symp Quant Biol. 1998. PMID: 10384325 Review. No abstract available.
Cited by
-
The neuronal host cell factor-binding protein Zhangfei inhibits herpes simplex virus replication.J Virol. 2005 Dec;79(23):14708-18. doi: 10.1128/JVI.79.23.14708-14718.2005. J Virol. 2005. PMID: 16282471 Free PMC article.
-
HDAC3 selectively represses CREB3-mediated transcription and migration of metastatic breast cancer cells.Cell Mol Life Sci. 2010 Oct;67(20):3499-510. doi: 10.1007/s00018-010-0388-5. Epub 2010 May 15. Cell Mol Life Sci. 2010. PMID: 20473547 Free PMC article.
-
A novel protein, Luman/CREB3 recruitment factor, inhibits Luman activation of the unfolded protein response.Mol Cell Biol. 2008 Jun;28(12):3952-66. doi: 10.1128/MCB.01439-07. Epub 2008 Apr 7. Mol Cell Biol. 2008. PMID: 18391022 Free PMC article.
-
HCF-1 functions as a coactivator for the zinc finger protein Krox20.J Biol Chem. 2003 Dec 19;278(51):51116-24. doi: 10.1074/jbc.M303470200. Epub 2003 Oct 6. J Biol Chem. 2003. PMID: 14532282 Free PMC article.
-
Molecular cloning of Drosophila HCF reveals proteolytic processing and self-association of the encoded protein.J Cell Physiol. 2003 Feb;194(2):117-26. doi: 10.1002/jcp.10193. J Cell Physiol. 2003. PMID: 12494450 Free PMC article.
References
-
- Flint J, Shenk T. Annu Rev Genet. 1997;31:177–212. - PubMed
-
- Weir J P. Gene. 2001;271:117–130. - PubMed
-
- O'Hare P. Semin Virol. 1993;4:145–155.
-
- Herr W. Cold Spring Harbor Symp Quant Biol. 1998;63:599–607. - PubMed
-
- Wilson A C, Cleary M A, Lai J S, LaMarco K, Peterson M G, Herr W. Cold Spring Harbor Symp Quant Biol. 1993;58:167–178. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
