

doi: 10.1101/gad.1013502.
A requirement for replication in activation of the ATR-dependent DNA damage checkpoint
Affiliations
- PMID: 12231621
- PMCID: PMC187437
- DOI: 10.1101/gad.1013502
Item in Clipboard
A requirement for replication in activation of the ATR-dependent DNA damage checkpoint
Genes Dev.
.
Abstract
Using the Xenopus egg extract system, we investigated the involvement of DNA replication in activation of the DNA damage checkpoint. We show here that DNA damage slows replication in a checkpoint-independent manner and is accompanied by replication-dependent recruitment of ATR and Rad1 to chromatin. We also find that the replication proteins RPA and Polalpha accumulate on chromatin following DNA damage. Finally, damage-induced Chk1 phosphorylation and checkpoint arrest are abrogated when replication is inhibited. These data indicate that replication is required for activation of the DNA damage checkpoint and suggest a unifying model for ATR activation by diverse lesions during S phase.
Figures
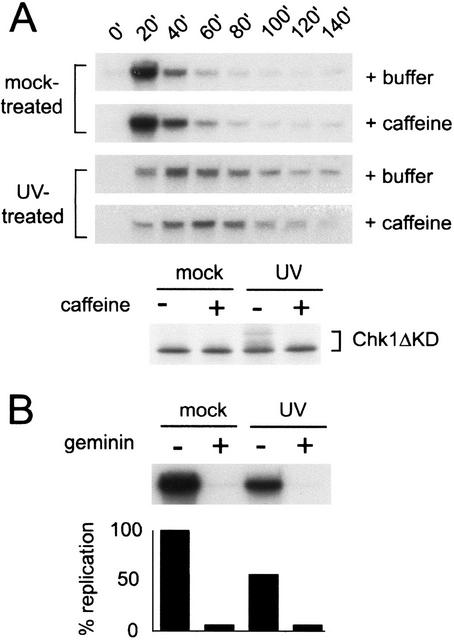
Replication is slowed in response to UV treatment. (A) Sperm nuclei (2000/μL) were mock-treated or UV-treated and added to Xenopus interphase extract in the presence (+caffeine) or absence (+buffer) of caffeine (4 mM), and the extract was divided into two samples. To assay replication, aliquots were removed from one sample at the given times, incubated with [α-32P]dCTP for 15 min, terminated, separated on a 0.8% agarose gel, and analyzed by autoradiography. To assay phosphorylation of xChk1, an in vitro translated, [35S]methionine-labeled fragment of xChk1 (Chk1ΔKD) was added to the second sample (5% reaction volume). Nuclei were isolated from this sample at 100 min, then proteins were separated by SDS-PAGE and analyzed by autoradiography. (B) CSF extract (10 μL) was preincubated with recombinant geminin or an equal volume of buffer. The extract was then supplemented with [α-32P]dCTP and mock- or UV-treated sperm nuclei, incubated at room temperature for 100 min, and processed as described in A.
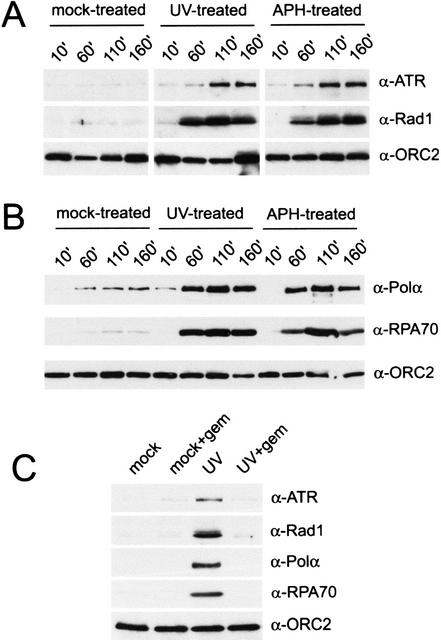
Geminin blocks UV-induced accumulation of checkpoint and replication proteins on chromatin. (A) Binding of xATR, xRad1, and xORC2 to chromatin in interphase extract was assayed after mock, UV, or aphidicolin (APH) treatment. Chromatin (2000/μL) was isolated at the given time points, and chromatin-bound proteins were analyzed by Western blotting with antibodies to xATR, xRad1, or xORC2. (B) Chromatin binding of the p180 subunit of xPolα, xRPA70, and xORC2 was assayed after mock, UV, and aphidicolin treatment by Western blotting. Samples were treated and prepared as described in A. (C) Interphase extract was pretreated for 10 min with geminin (+gem) before addition of mock- or UV-treated chromatin (2000/μL). Chromatin was isolated after 100 min, and chromatin-bound proteins were analyzed as described above.
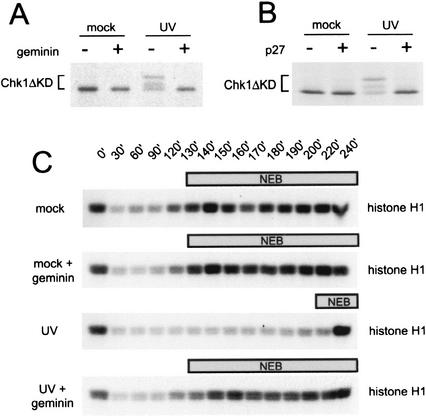
Loss of the DNA damage checkpoint response after inhibition of DNA replication. (A) Phosphorylation of xChk1 (Chk1ΔKD) in Xenopus interphase extract after geminin treatment. Mock-treated or UV-damaged sperm chromatin (2000/μL) was added to mock- or geminin-pretreated extract containing Chk1ΔKD, and samples were analyzed as described in Figure 1A. (B) Phosphorylation of xChk1 (Chk1ΔKD) in Xenopus interphase extract after p27 treatment. Mock-treated or UV-damaged sperm chromatin (2000/μL) was added to mock- or p27-pretreated extract containing Chk1ΔKD, and samples were analyzed as described in Figure 1A. (C) Phosphorylation of histone H1 in mock- or geminin-treated cytostatic-factor-arrested (CSF) extract. CSF extract pretreated with buffer or recombinant geminin was supplemented with mock- or UV-treated sperm nuclei and 1 mM CaCl2. At the indicated time points, samples were removed to assay histone H1 kinase activity. The time at which nuclear envelope breakdown (NEB) was >50% is indicated by the bar above each panel.
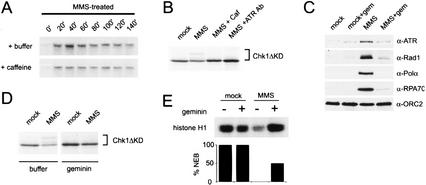
Activation of the MMS checkpoint is replication-dependent. (A) DNA replication is slowed after MMS damage. Chromatin was pretreated with MMS, and replication was assayed as described in Figure 1A. See Figure 1A for untreated control (+/−) caffeine. (B) Phosphorylation of xChk1ΔKD induced by MMS was assayed after treatment of the extract with the checkpoint kinase inhibitor caffeine (4 mM) or an xATR-neutralizing antibody (10% reaction volume) as described in Figure 1A. (C) Chromatin was isolated from mock- or geminin-treated extract containing mock- or MMS-treated chromatin after 100 min, and chromatin-bound proteins were analyzed as described in Figure 2. (D) MMS-induced phosphorylation of xChk1ΔKD was assayed after geminin treatment. Geminin-treated interphase extract was supplemented with xChk1ΔKD and sperm chromatin (3000/μL), and samples were harvested at 100 min. Sperm nuclei were treated with MMS prior to addition to the extract. (E) Histone H1 kinase activity and percentage of nuclear envelope breakdown (%NEB) were assayed in CSF extract at 160 min. Extract was pretreated with buffer or geminin, and sperm chromatin was mock-treated or treated with MMS prior to addition to extract.
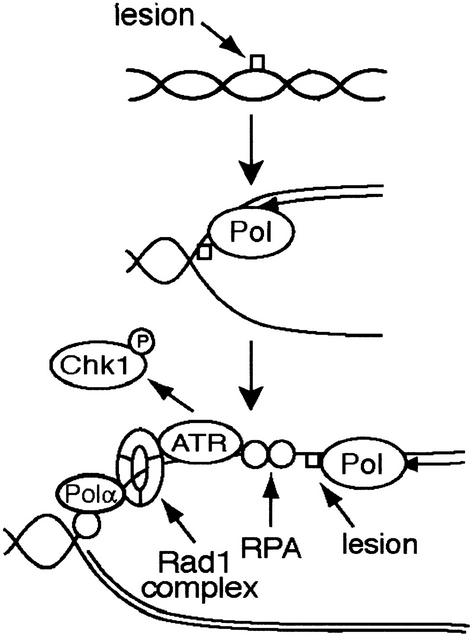
A model for activation of the DNA damage checkpoint in Xenopus egg extracts. UV and MMS lesions do not activate the checkpoint prior to initiation of DNA replication. During DNA replication, we suggest that encounter of a polymerase with a lesion leads to slowing of replication. Accumulation of single-stranded DNA is followed by association of ATR and the Rad1 complex with a protein and/or nucleic acid structure that is dependent on the presence of RPA, Polα, and possibly additional downstream events. Activation of ATR and Chk1 phosphorylation follow.
Similar articles
-
Requirement for Atr in phosphorylation of Chk1 and cell cycle regulation in response to DNA replication blocks and UV-damaged DNA in Xenopus egg extracts.Genes Dev. 2000 Nov 1;14(21):2745-56. doi: 10.1101/gad.842500. Genes Dev. 2000. PMID: 11069891 Free PMC article.
-
Xenopus ATR is a replication-dependent chromatin-binding protein required for the DNA replication checkpoint.Curr Biol. 2000 Dec 14-28;10(24):1565-73. doi: 10.1016/s0960-9822(00)00855-1. Curr Biol. 2000. PMID: 11137007
-
Claspin, a Chk1-regulatory protein, monitors DNA replication on chromatin independently of RPA, ATR, and Rad17.Mol Cell. 2003 Feb;11(2):329-40. doi: 10.1016/s1097-2765(03)00045-5. Mol Cell. 2003. PMID: 12620222
-
Chk1 in the DNA damage response: conserved roles from yeasts to mammals.DNA Repair (Amst). 2004 Aug-Sep;3(8-9):1025-32. doi: 10.1016/j.dnarep.2004.03.003. DNA Repair (Amst). 2004. PMID: 15279789 Review.
-
Safeguarding genome integrity: the checkpoint kinases ATR, CHK1 and WEE1 restrain CDK activity during normal DNA replication.Nucleic Acids Res. 2012 Jan;40(2):477-86. doi: 10.1093/nar/gkr697. Epub 2011 Sep 21. Nucleic Acids Res. 2012. PMID: 21937510 Free PMC article. Review.
Cited by
-
An ATR- and BRCA1-mediated Fanconi anemia pathway is required for activating the G2/M checkpoint and DNA damage repair upon rereplication.Mol Cell Biol. 2006 Jun;26(12):4601-11. doi: 10.1128/MCB.02141-05. Mol Cell Biol. 2006. PMID: 16738325 Free PMC article.
-
Minichromosome maintenance proteins are direct targets of the ATM and ATR checkpoint kinases.Proc Natl Acad Sci U S A. 2004 Jul 6;101(27):10078-83. doi: 10.1073/pnas.0403410101. Epub 2004 Jun 21. Proc Natl Acad Sci U S A. 2004. PMID: 15210935 Free PMC article.
-
Molecular mechanisms of DNA replication checkpoint activation.Genes (Basel). 2014 Mar 6;5(1):147-75. doi: 10.3390/genes5010147. Genes (Basel). 2014. PMID: 24705291 Free PMC article.
-
MMB-FOXM1-driven premature mitosis is required for CHK1 inhibitor sensitivity.Cell Rep. 2021 Mar 2;34(9):108808. doi: 10.1016/j.celrep.2021.108808. Cell Rep. 2021. PMID: 33657372 Free PMC article.
-
Continued primer synthesis at stalled replication forks contributes to checkpoint activation.J Cell Biol. 2010 Apr 19;189(2):233-46. doi: 10.1083/jcb.200909105. Epub 2010 Apr 12. J Cell Biol. 2010. PMID: 20385778 Free PMC article.
References
-
- Abraham RT. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes & Dev. 2001;15:2177–2196. - PubMed
-
- Cortez D, Guntuku S, Qin J, Elledge SJ. ATR and ATRIP: Partners in checkpoint signaling. Science. 2001;294:1713–1716. - PubMed
-
- D'Urso G, Grallert B, Nurse P. DNA polymerase α, a component of the replication initiation complex, is essential for the checkpoint coupling S phase to mitosis in fission yeast. J Cell Sci. 1995;108:3109–3118. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous