

Brain factor-1 controls the proliferation and differentiation of neocortical progenitor cells through independent mechanisms
- PMID: 12151532
- PMCID: PMC6758167
- DOI: 10.1523/JNEUROSCI.22-15-06526.2002
Brain factor-1 controls the proliferation and differentiation of neocortical progenitor cells through independent mechanisms
Abstract
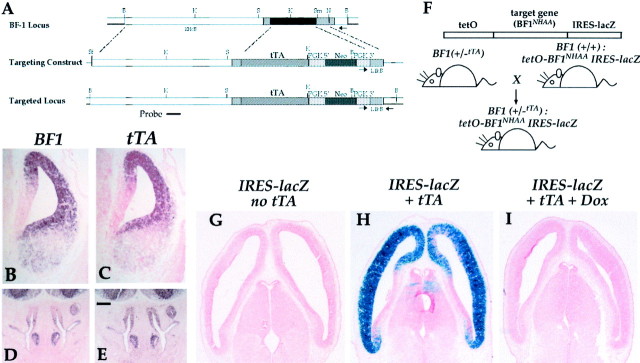
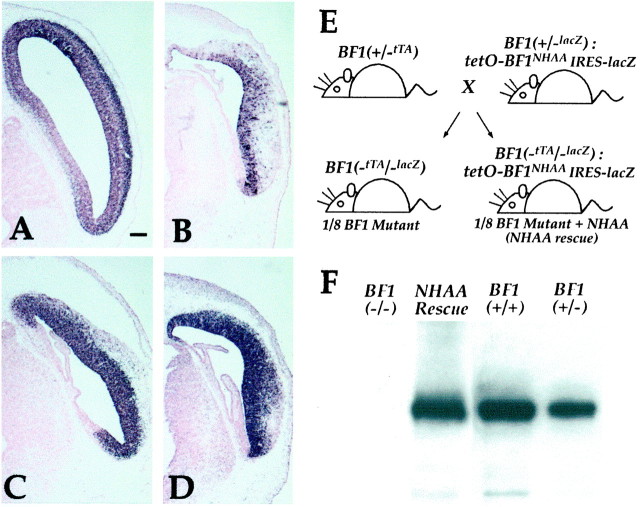
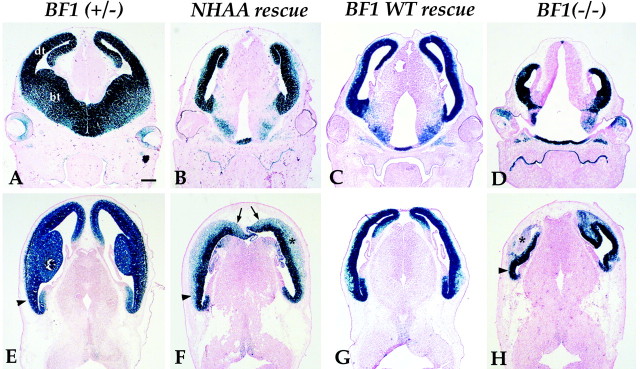
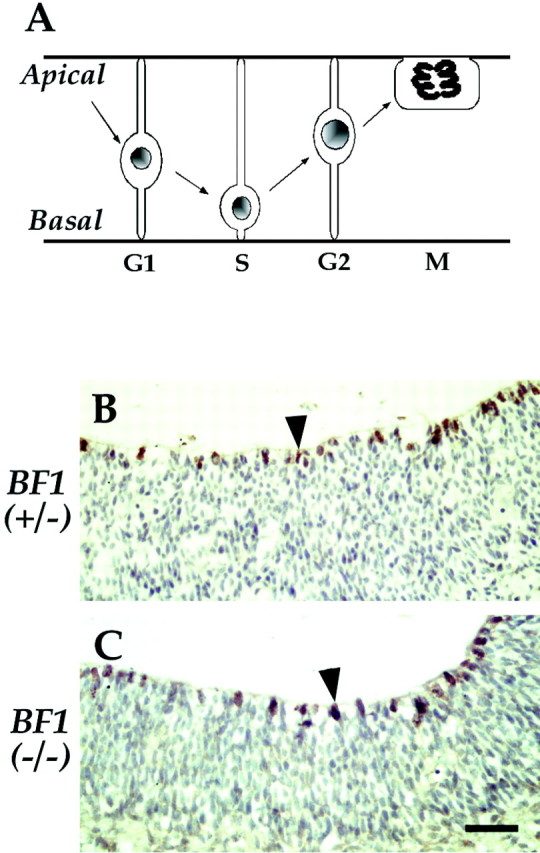
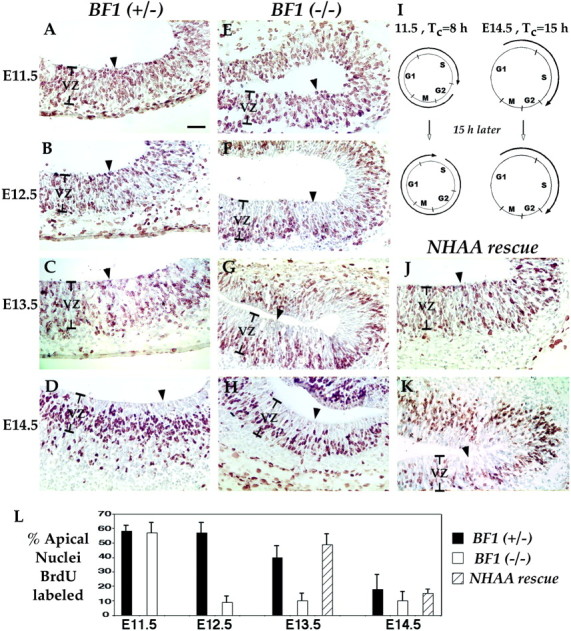
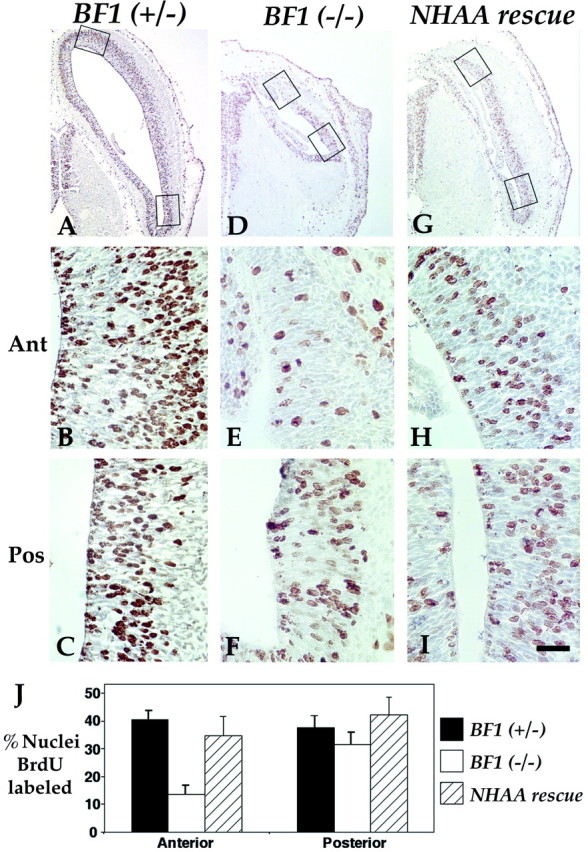
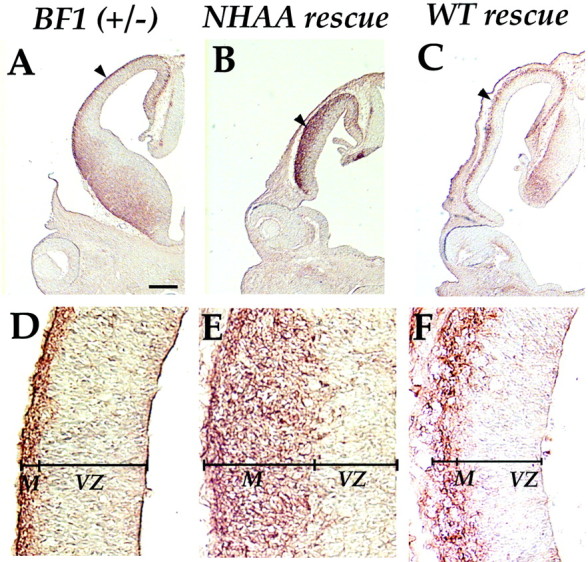
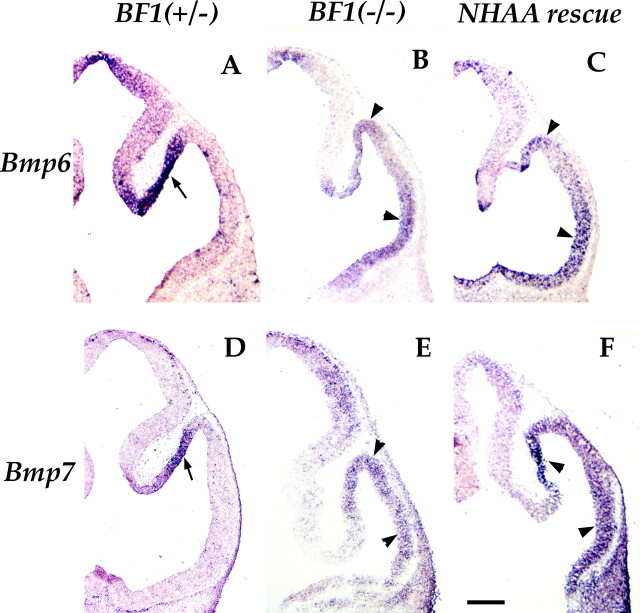
The winged helix gene Brain factor-1 (BF1) has a pleiotropic role in the development of the cerebral hemispheres of the brain. Mice lacking BF1 have defects in the morphogenesis of the structures of the dorsal telencephalon (e.g., neocortex) and the ventral telencephalon (e.g., the basal ganglia). This study focuses on the functions of BF1 in the dorsal telencephalon. We showed previously that telencephalic progenitor cells lacking BF1 differentiate into neurons prematurely. Here, we demonstrate that the loss of BF1 also results in an early lengthening of the cell cycle in neocortical progenitors. To investigate the mechanisms by which BF1 regulates progenitor cell proliferation and differentiation in the developing brain, we have replaced the endogenous BF1 protein with a DNA binding defective form of BF1 in mice, BF1(NHAA). The BF1(NHAA) protein restores the growth of the dorsal telencephalon, by improving the proliferation of progenitor cells. However, the BF1(NHAA) protein does not correct the early neuronal differentiation associated with the loss of BF1. In contrast, replacement of endogenous BF1 with wild-type BF1 corrects the defects in both the proliferation and differentiation of neocortical progenitors. These results demonstrate that BF1 controls progenitor cell proliferation and differentiation in the neocortex through distinct DNA binding-independent and binding-dependent mechanisms.
Figures
Similar articles
-
Dual role of brain factor-1 in regulating growth and patterning of the cerebral hemispheres.Cereb Cortex. 1999 Sep;9(6):543-50. doi: 10.1093/cercor/9.6.543. Cereb Cortex. 1999. PMID: 10498272
-
Winged helix transcription factor BF-1 is essential for the development of the cerebral hemispheres.Neuron. 1995 Jun;14(6):1141-52. doi: 10.1016/0896-6273(95)90262-7. Neuron. 1995. PMID: 7605629
-
Foxg1 is required for specification of ventral telencephalon and region-specific regulation of dorsal telencephalic precursor proliferation and apoptosis.Dev Biol. 2005 Jul 1;283(1):113-27. doi: 10.1016/j.ydbio.2005.04.005. Dev Biol. 2005. PMID: 15893304
-
The progenitor cells of the embryonic telencephalon and the neonatal anterior subventricular zone differentially regulate their cell cycle.Chem Senses. 2002 Jul;27(6):577-80. doi: 10.1093/chemse/27.6.577. Chem Senses. 2002. PMID: 12142335 Free PMC article. Review.
-
Sonic hedgehog signaling coordinates the proliferation and differentiation of neural stem/progenitor cells by regulating cell cycle kinetics during development of the neocortex.Congenit Anom (Kyoto). 2012 Jun;52(2):72-7. doi: 10.1111/j.1741-4520.2012.00368.x. Congenit Anom (Kyoto). 2012. PMID: 22639991 Review.
Cited by
-
Haploinsufficiency of novel FOXG1B variants in a patient with severe mental retardation, brain malformations and microcephaly.Hum Genet. 2005 Oct;117(6):536-44. doi: 10.1007/s00439-005-1310-3. Epub 2005 Aug 17. Hum Genet. 2005. PMID: 16133170
-
Antagonistic effects of Grg6 and Groucho/TLE on the transcription repression activity of brain factor 1/FoxG1 and cortical neuron differentiation.Mol Cell Biol. 2005 Dec;25(24):10916-29. doi: 10.1128/MCB.25.24.10916-10929.2005. Mol Cell Biol. 2005. PMID: 16314515 Free PMC article.
-
Foxg1 haploinsufficiency reduces the population of cortical intermediate progenitor cells: effect of increased p21 expression.Cereb Cortex. 2008 Aug;18(8):1865-75. doi: 10.1093/cercor/bhm209. Epub 2007 Dec 7. Cereb Cortex. 2008. PMID: 18065723 Free PMC article.
-
Molecular control of neurogenesis: a view from the mammalian cerebral cortex.Cold Spring Harb Perspect Biol. 2012 Oct 1;4(10):a008359. doi: 10.1101/cshperspect.a008359. Cold Spring Harb Perspect Biol. 2012. PMID: 23028117 Free PMC article. Review.
-
FOXG1 dysregulation is a frequent event in medulloblastoma.J Neurooncol. 2007 Nov;85(2):111-22. doi: 10.1007/s11060-007-9394-3. Epub 2007 May 24. J Neurooncol. 2007. PMID: 17522785
References
-
- Bourguignon C, Li J, Papalopulu N. XBF-1, a winged helix transcription factor with dual activity, has a role in positioning neurogenesis in Xenopus competent ectoderm. Development. 1998;125:4889–4900. - PubMed
-
- Caviness VJ, Takahashi T, Nowakowski R. Numbers, time and neocortical neuronogenesis: a general developmental and evolutionary model. Trends Neurosci. 1995;18:379–383. - PubMed
-
- Clark D, Mitra P, Wang S. Scalable architecture in mammalian brains. Nature. 2001;411:189–193. - PubMed
-
- Dewulf N, Verschueren K, Lonnoy O, Moren A, Grimsby S, Vande Spiegle K, Miyazono K, Hulebroeck D, ten Dijke P. Distinct spatial and temporal expression patterns of two type I receptors for bone morphogenetic proteins during mouse embryogenesis. Endocrinology. 1995;136:2652–2663. - PubMed
-
- Dou C, Li S, Lai E. Dual role of brain factor-1 in regulating growth and patterning of the cerebral hemispheres. Cereb Cortex. 1999;9:543–550. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases