

A neuronal glutamate transporter contributes to neurotransmitter GABA synthesis and epilepsy
- PMID: 12151515
- PMCID: PMC2483507
- DOI: 10.1523/JNEUROSCI.22-15-06372.2002
A neuronal glutamate transporter contributes to neurotransmitter GABA synthesis and epilepsy
Abstract
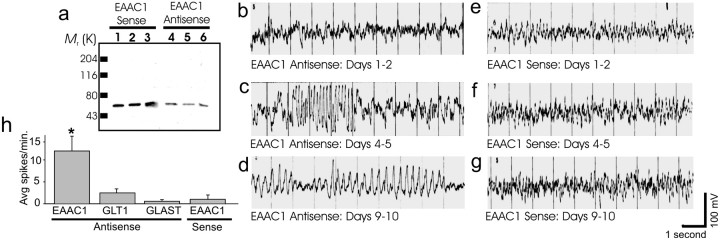
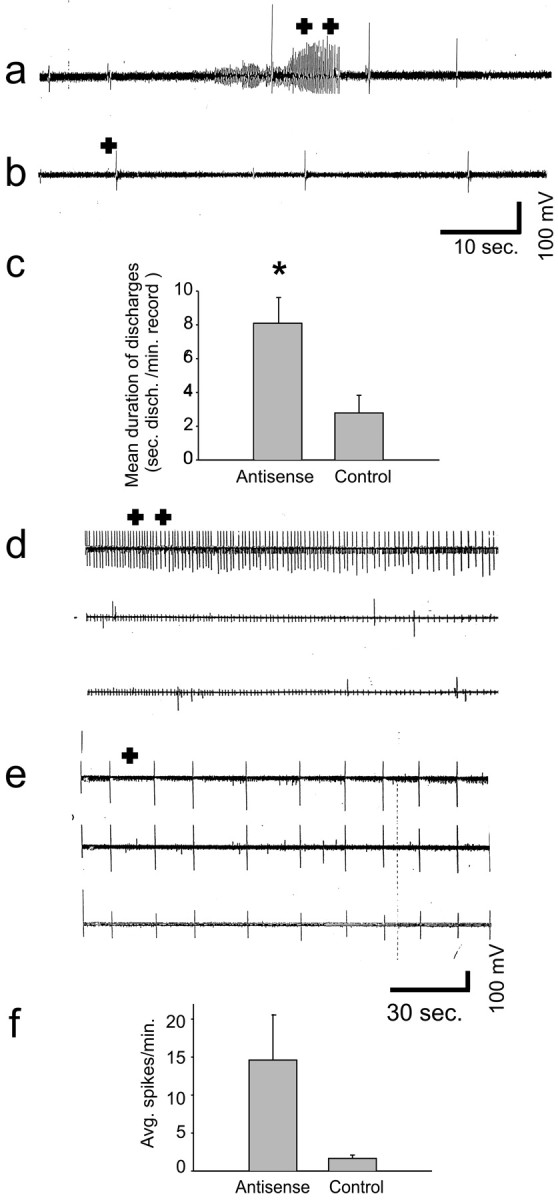
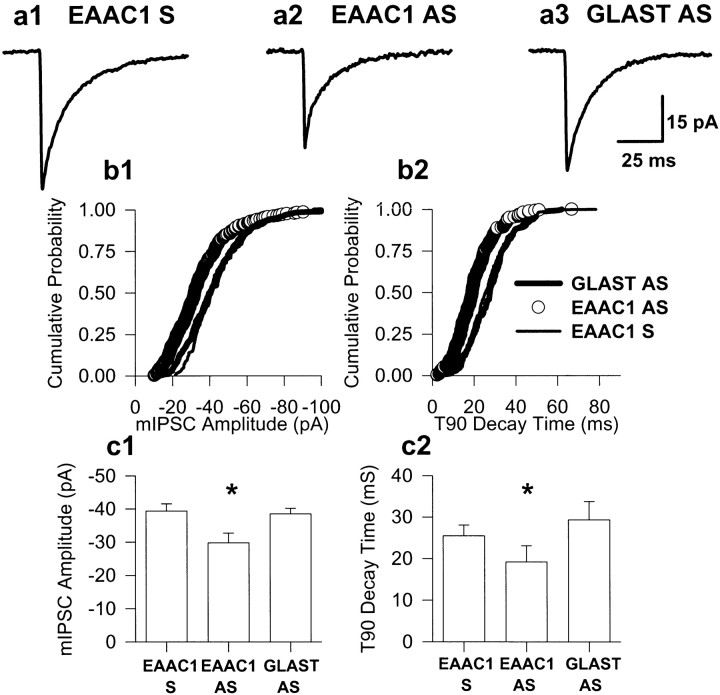
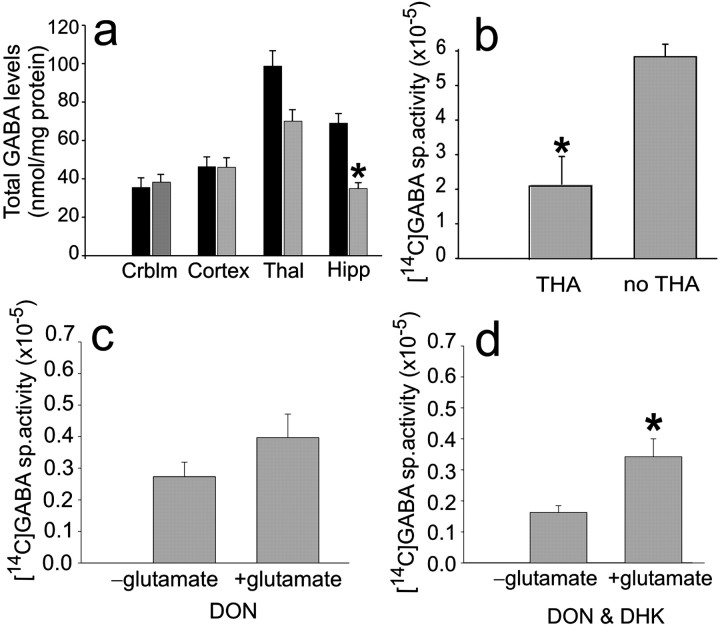
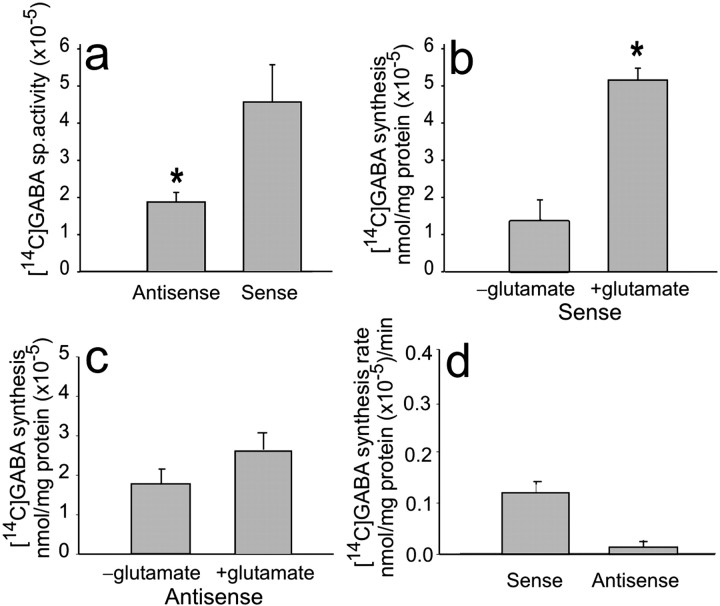
The predominant neuronal glutamate transporter, EAAC1 (for excitatory amino acid carrier-1), is localized to the dendrites and somata of many neurons. Rare presynaptic localization is restricted to GABA terminals. Because glutamate is a precursor for GABA synthesis, we hypothesized that EAAC1 may play a role in regulating GABA synthesis and, thus, could cause epilepsy in rats when inactivated. Reduced expression of EAAC1 by antisense treatment led to behavioral abnormalities, including staring-freezing episodes and electrographic (EEG) seizures. Extracellular hippocampal and thalamocortical slice recordings showed excessive excitability in antisense-treated rats. Patch-clamp recordings of miniature IPSCs (mIPSCs) conducted in CA1 pyramidal neurons in slices from EAAC1 antisense-treated animals demonstrated a significant decrease in mIPSC amplitude, indicating decreased tonic inhibition. There was a 50% loss of hippocampal GABA levels associated with knockdown of EAAC1, and newly synthesized GABA from extracellular glutamate was significantly impaired by reduction of EAAC1 expression. EAAC1 may participate in normal GABA neurosynthesis and limbic hyperexcitability, whereas epilepsy can result from a disruption of the interaction between EAAC1 and GABA metabolism.
Figures
Similar articles
-
Translocation of glutamate transporter subtype excitatory amino acid carrier 1 protein in kainic acid-induced rat epilepsy.Am J Pathol. 2003 Aug;163(2):779-87. doi: 10.1016/S0002-9440(10)63705-4. Am J Pathol. 2003. PMID: 12875997 Free PMC article.
-
Glutamate transporters alterations in the reorganizing dentate gyrus are associated with progressive seizure activity in chronic epileptic rats.J Comp Neurol. 2002 Jan 21;442(4):365-77. doi: 10.1002/cne.10101. J Comp Neurol. 2002. PMID: 11793340
-
Neuronal glutamate uptake Contributes to GABA synthesis and inhibitory synaptic strength.J Neurosci. 2003 Mar 15;23(6):2040-8. doi: 10.1523/JNEUROSCI.23-06-02040.2003. J Neurosci. 2003. PMID: 12657662 Free PMC article.
-
Chronic hyperammonemia alters extracellular glutamate, glutamine and GABA and membrane expression of their transporters in rat cerebellum. Modulation by extracellular cGMP.Neuropharmacology. 2019 Dec 15;161:107496. doi: 10.1016/j.neuropharm.2019.01.011. Epub 2019 Jan 11. Neuropharmacology. 2019. PMID: 30641078 Review.
-
Changes in the expression of the glutamate transporter EAAT3/EAAC1 in health and disease.Cell Mol Life Sci. 2014 Jun;71(11):2001-15. doi: 10.1007/s00018-013-1484-0. Epub 2013 Oct 26. Cell Mol Life Sci. 2014. PMID: 24162932 Free PMC article. Review.
Cited by
-
Functional studies and rare variant screening of SLC1A1/EAAC1 in males with obsessive-compulsive disorder.Psychiatr Genet. 2012 Oct;22(5):256-60. doi: 10.1097/YPG.0b013e328353fb63. Psychiatr Genet. 2012. PMID: 22617815 Free PMC article.
-
Parasynaptic NMDA receptor signaling couples neuronal glutamate transporter function to AMPA receptor synaptic distribution and stability.J Neurosci. 2012 Feb 15;32(7):2552-63. doi: 10.1523/JNEUROSCI.3237-11.2012. J Neurosci. 2012. PMID: 22396428 Free PMC article.
-
Glutamate Spillover Dynamically Strengthens Gabaergic Synaptic Inhibition of the Hypothalamic Paraventricular Nucleus.J Neurosci. 2024 Feb 14;44(7):e1851222023. doi: 10.1523/JNEUROSCI.1851-22.2023. J Neurosci. 2024. PMID: 38154957 Free PMC article.
-
Rapid compensatory changes in the expression of EAAT-3 and GAT-1 transporters during seizures in cells of the CA1 and dentate gyrus.J Biomed Sci. 2012 Aug 29;19(1):78. doi: 10.1186/1423-0127-19-78. J Biomed Sci. 2012. PMID: 22931236 Free PMC article.
-
A computational model to investigate astrocytic glutamate uptake influence on synaptic transmission and neuronal spiking.Front Comput Neurosci. 2012 Oct 1;6:70. doi: 10.3389/fncom.2012.00070. eCollection 2012. Front Comput Neurosci. 2012. PMID: 23060782 Free PMC article.
References
-
- Agmon A, Connors BW. Thalamocortical responses of mouse somatosensory (barrel) cortex in vitro. Neuroscience. 1991;41:365–379. - PubMed
-
- Behar KL, Boehm D. Measurement of GABA following GABA-transaminase inhibition by gabaculine: a 1H and 31P NMR spectroscopic study of rat brain in vivo. Magn Reson Med. 1994;31:660–667. - PubMed
-
- Bergles DE, Jahr CE. Synaptic activation of glutamate transporters in hippocampal astrocytes. Neuron. 1997;19:1297–1308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS032403-09/NS/NINDS NIH HHS/United States
- R01 NS038572/NS/NINDS NIH HHS/United States
- R01 NS040151/NS/NINDS NIH HHS/United States
- R01 NS038572-04/NS/NINDS NIH HHS/United States
- NS 32403/NS/NINDS NIH HHS/United States
- R01 NS036465/NS/NINDS NIH HHS/United States
- R01 NS032403/NS/NINDS NIH HHS/United States
- R37 NS032403/NS/NINDS NIH HHS/United States
- NS33958/NS/NINDS NIH HHS/United States
- NS36465/NS/NINDS NIH HHS/United States
- NS 38572/NS/NINDS NIH HHS/United States
- NS 40151/NS/NINDS NIH HHS/United States
- R01 NS033958/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous