

Identification of a family of calcium sensors as protein ligands of inositol trisphosphate receptor Ca(2+) release channels
- PMID: 12032348
- PMCID: PMC124329
- DOI: 10.1073/pnas.102006299
Identification of a family of calcium sensors as protein ligands of inositol trisphosphate receptor Ca(2+) release channels
Abstract
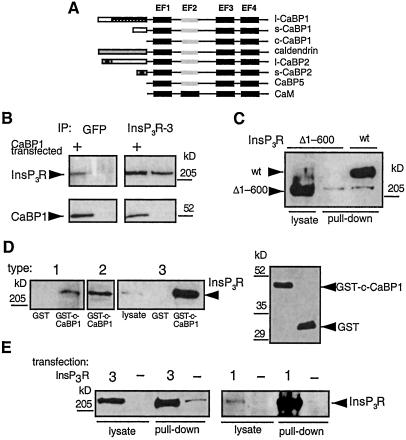
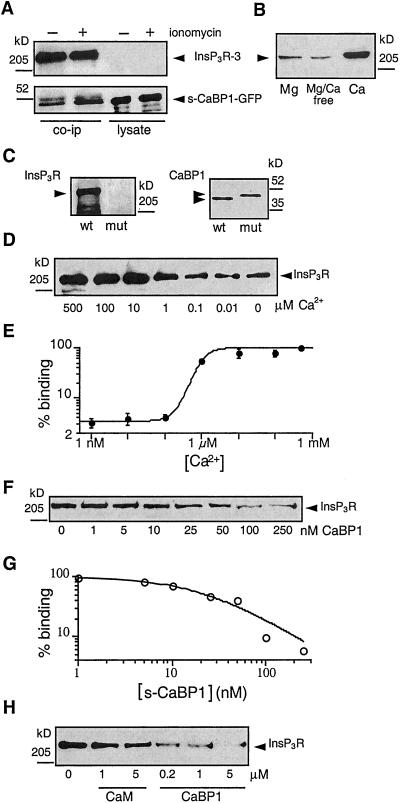
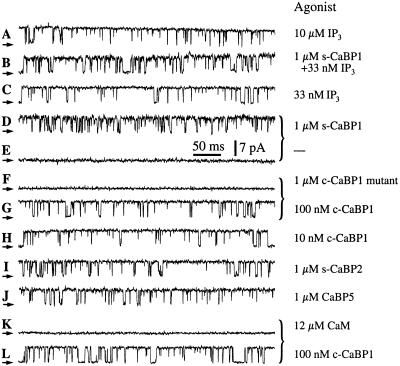
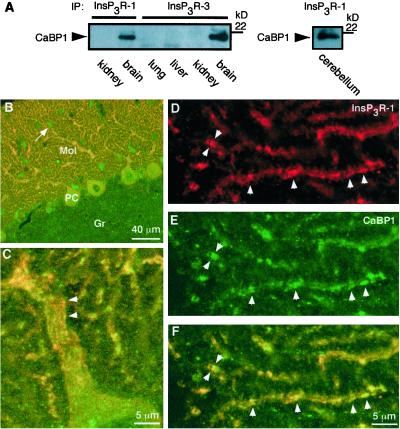
The inositol trisphosphate (InsP(3)) receptor (InsP(3)R) is a ubiquitously expressed intracellular Ca(2+) channel that mediates complex cytoplasmic Ca(2+) signals, regulating diverse cellular processes, including synaptic plasticity. Activation of the InsP(3)R channel is normally thought to require binding of InsP(3) derived from receptor-mediated activation of phosphatidylinositol lipid hydrolysis. Here we identify a family of neuronal Ca(2+)-binding proteins as high-affinity protein agonists of the InsP(3)R, which bind to the channel and activate gating in the absence of InsP(3). CaBP/caldendrin, a subfamily of the EF-hand-containing neuronal calcium sensor family of calmodulin-related proteins, bind specifically to the InsP(3)-binding region of all three InsP(3)R channel isoforms with high affinity (K(a) approximately 25 nM) in a Ca(2+)-dependent manner (K(a) approximately 1 microM). Binding activates single-channel gating as efficaciously as InsP(3), dependent on functional EF-hands in CaBP. In contrast, calmodulin neither bound with high affinity nor activated channel gating. CaBP1 and the type 1 InsP(3)R associate in rat whole brain and cerebellum lysates, and colocalize extensively in subcellular regions in cerebellar Purkinje neurons. Thus, InsP(3)R-mediated Ca(2+) signaling in cells is possible even in the absence of InsP(3) generation, a process that may be particularly important in responding to and shaping changes in intracellular Ca(2+) concentration by InsP(3)-independent pathways and for localizing InsP(3)-mediated Ca(2+) signals to individual synapses.
Figures
Comment in
-
Activating calcium release through inositol 1,4,5-trisphosphate receptors without inositol 1,4,5-trisphosphate.Proc Natl Acad Sci U S A. 2002 May 28;99(11):7320-2. doi: 10.1073/pnas.132254299. Proc Natl Acad Sci U S A. 2002. PMID: 12032280 Free PMC article. Review. No abstract available.
Similar articles
-
Single-channel properties in endoplasmic reticulum membrane of recombinant type 3 inositol trisphosphate receptor.J Gen Physiol. 2000 Mar;115(3):241-56. doi: 10.1085/jgp.115.3.241. J Gen Physiol. 2000. PMID: 10694253 Free PMC article.
-
ATP-dependent adenophostin activation of inositol 1,4,5-trisphosphate receptor channel gating: kinetic implications for the durations of calcium puffs in cells.J Gen Physiol. 2001 Apr;117(4):299-314. doi: 10.1085/jgp.117.4.299. J Gen Physiol. 2001. PMID: 11279251 Free PMC article.
-
CIB1, a ubiquitously expressed Ca2+-binding protein ligand of the InsP3 receptor Ca2+ release channel.J Biol Chem. 2006 Jul 28;281(30):20825-20833. doi: 10.1074/jbc.M602175200. Epub 2006 May 24. J Biol Chem. 2006. PMID: 16723353
-
Nuclear Ca2+ signalling in cerebellar Purkinje neurons.Cerebellum. 2006;5(1):36-42. doi: 10.1080/14734220600554438. Cerebellum. 2006. PMID: 16527762 Review.
-
Modulation of Ca2+ oscillations by phosphorylation of Ins(1,4,5)P3 receptors.Biochem Soc Trans. 2003 Oct;31(Pt 5):954-7. doi: 10.1042/bst0310954. Biochem Soc Trans. 2003. PMID: 14505457 Review.
Cited by
-
Activating calcium release through inositol 1,4,5-trisphosphate receptors without inositol 1,4,5-trisphosphate.Proc Natl Acad Sci U S A. 2002 May 28;99(11):7320-2. doi: 10.1073/pnas.132254299. Proc Natl Acad Sci U S A. 2002. PMID: 12032280 Free PMC article. Review. No abstract available.
-
Thimerosal stimulates Ca2+ flux through inositol 1,4,5-trisphosphate receptor type 1, but not type 3, via modulation of an isoform-specific Ca2+-dependent intramolecular interaction.Biochem J. 2004 Jul 1;381(Pt 1):87-96. doi: 10.1042/BJ20040072. Biochem J. 2004. PMID: 15015936 Free PMC article.
-
Identification and functional distribution of intracellular ca channels in mouse lacrimal gland acinar cells.Open Ophthalmol J. 2007 Dec 4;1:8-16. doi: 10.2174/1874364100701010008. Open Ophthalmol J. 2007. PMID: 19478858 Free PMC article.
-
Protein phosphatase 2A dephosphorylates CaBP4 and regulates CaBP4 function.Invest Ophthalmol Vis Sci. 2013 Feb 1;54(2):1214-26. doi: 10.1167/iovs.12-11319. Invest Ophthalmol Vis Sci. 2013. PMID: 23341017 Free PMC article.
-
Complicity of α-synuclein oligomer and calcium dyshomeostasis in selective neuronal vulnerability in Lewy body disease.Arch Pharm Res. 2021 Jun;44(6):564-573. doi: 10.1007/s12272-021-01334-6. Epub 2021 Jun 10. Arch Pharm Res. 2021. PMID: 34114191 Free PMC article. Review.
References
-
- Berridge M J. Nature (London) 1993;361:315–325. - PubMed
-
- Berridge M J. J Exp Biol. 1997;200:315–319. - PubMed
-
- Berridge M J, Bootman M D, Lipp P. Nature (London) 1998;395:645–648. - PubMed
-
- Taylor C W, Genazzani A A, Morris S A. Cell Calcium. 1999;26:237–251. - PubMed
-
- Patel S, Joseph S K, Thomas A P. Cell Calcium. 1999;25:247–264. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
