

doi: 10.1073/pnas.112212199.
Disruption of a long-range cis-acting regulator for Shh causes preaxial polydactyly
Affiliations
- PMID: 12032320
- PMCID: PMC124279
- DOI: 10.1073/pnas.112212199
Item in Clipboard
Disruption of a long-range cis-acting regulator for Shh causes preaxial polydactyly
Proc Natl Acad Sci U S A.
.
Abstract
Preaxial polydactyly (PPD) is a common limb malformation in human. A number of polydactylous mouse mutants indicate that misexpression of Shh is a common requirement for generating extra digits. Here we identify a translocation breakpoint in a PPD patient and a transgenic insertion site in the polydactylous mouse mutant sasquatch (Ssq). The genetic lesions in both lie within the same respective intron of the LMBR1/Lmbr1 gene, which resides approximately 1 Mb away from Shh. Genetic analysis of Ssq reveals that the Lmbr1 gene is incidental to the phenotype and that the mutation directly interrupts a cis-acting regulator of Shh. This regulator is most likely the target for generating PPD mutations in human.
Figures
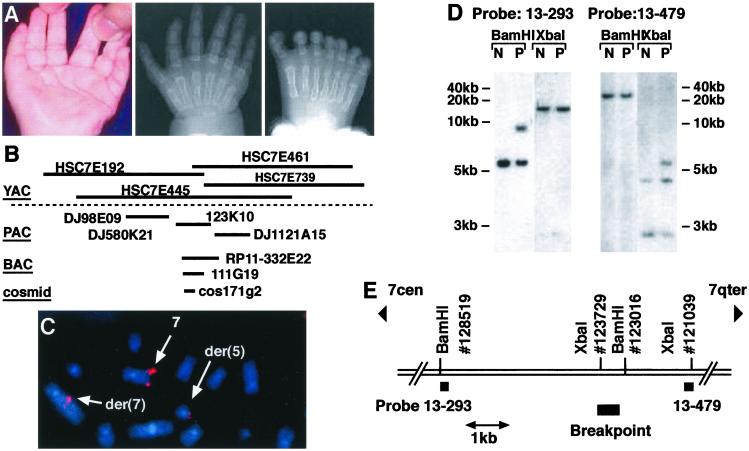
Phenotype and identification of the breakpoint in the t(5 7)(q11,q36)translocation patient. (A) Left hand (Left, ventral view) of the PPD translocation patient showing duplication of the triphalangeal thumb. Radiogram of the right hand (Center) showing bilateral duplications and of the right foot (Right) showing triplication of the great toe. (B) The YACs, PACs, BACs, and a cosmid used to define the translocation breakpoint. (C) FISH analysis of metaphase chromosomes from a lymphoblastoid cell line from the patient, using cosmid clone171G2 as a probe. Hybridization signals are marked by arrows on the wild-type chromosome 7 and on the products of the translocation, the derived chromosome 5 (der 5) and derived chromosome 7 (der 7). (D) Southern blot analysis with genomic DNA from the lymphoblastoid cell line from the patient (P) and a normal control (N). DNA was digested with restriction endonucleases BamHI and XbaI. STS probe 13–293 (Left) and STS probe 13–479 (Right) were used. The relative positions of the STS probes, restriction sites, and the translocation breakpoint are indicated (E).
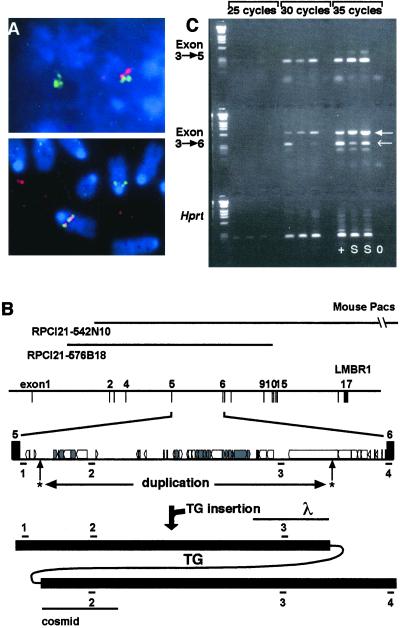
Identification of the mouse Ssq insertion site. PAC RPCI21-542N10 was isolated as containing the transgene integration site. (A) FISH analysis confirms the colocalization of the PAC with the transgene. PAC RPCI21-542N10 is labeled in green and the human placental alkaline phosphatase reporter gene (HPAP)-containing transgene is labeled in red. (Upper) Colocalization in an interphase nucleus; (Lower) colocalization in a metaphase spread. (B) The regions of the Lmbr1 gene covered by the two PAC clones (RPCI21 542N10 and 576B18) isolated. The position of the intron in the Lmbr1 gene (exon/intron gene structure from human sequence) and the intronic repeat structure (boxes) derived from the complete sequence between exons 5 and 6 is shown. Unique sequence DNA was predicted and four probes were generated (positions shown below intronic structure numbered 1–4). The positions of transgene integration (arrows with asterisks) and the area of intron duplication (below asterisks) are indicated. The resolved structure after integration is shown (bottom line), indicating intronic duplication as defined by probes 1–4. The positions of the λ and cosmid genomic clones (lines underneath) that defined the integration sites are marked. (C) RT-PCR samples were removed after 25, 30, and 35 cycles. RNA is from 11.5-day embryos of wild-type (+) and Ssq/Ssq (S) genotypes; a no RNA control (0) is also included. Top and Middle make use of primer pairs specific to Lmbr1; the first is from exon 3 to exon 5 and examines expression levels 5′ of the transgene integration. Comparable levels are detected in wild-type and Ssq/Ssq embryos. Middle uses primers that cross the integration site spanning exons 3 and 6. With these, at 30 cycles a product is only detected in the wild-type sample. The primers used in this experiment cross-hybridize with the mouse glycosylphosphatidylinositol-anchored protein homologue (closed arrowhead), which serves as positive control for the experiment and to normalize the samples. (Bottom) primers from the HPRT locus were used as positive control for RNA integrity and controls for reverse transcription.
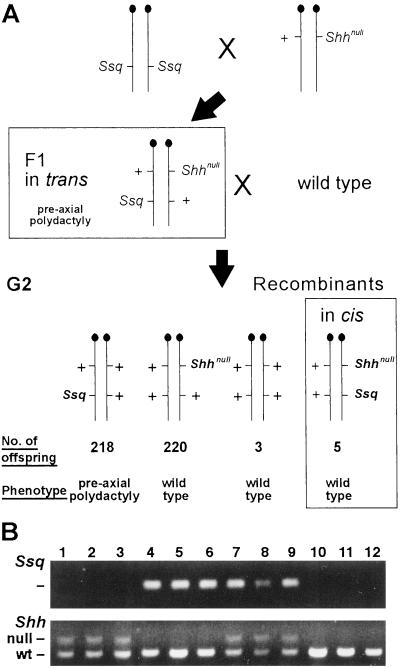
Cis–trans genetic test. A shows the genetic cross used. A homozygous Ssq mouse was crossed to a Shhnull heterozygote. The F1 mice that carry both mutations on opposite chromosomes, all of which exhibit preaxial polydactyly, were crossed to wild-type mice. The bottom row depicts the outcome of the cross. The majority of G2 progeny were, as expected, found to be heterozygous for either the Ssq allele or the Shhnull (218 and 220 mice, respectively). The mice of interest were those identified as carrying a chromosome 5 that has recombined between the Shh locus and the region of the Ssq insertion. Three had inherited the wild-type alleles at both loci and five carried the Ssq allele and the Shhnull allele in cis. These mice show no additional digits and have wild-type paws. This cross results in a genetic distance of 1.8 cM between Shh and Ssq. B shows examples of the genotyping PCRs used to analyze the G2 progeny. Lanes 1–3 are mice heterozygous for the Shhnull allele. Lanes 4–6 are wild type for Shh but carry the Ssq insertion. Mice in lanes 7–12 carry the recombinant chromosomes; 7–9 show G2s with both Shhnull and Ssq, whereas 10–12 are wild type at both loci.
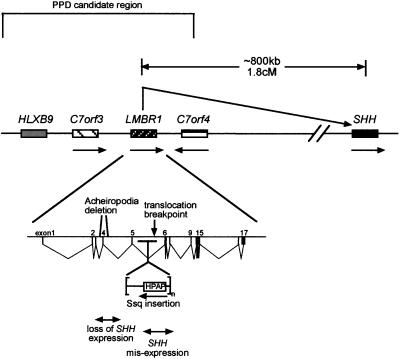
Diagram of the human PPD critical region on chromosome 7q36 and the relationship with Shh. The corresponding region in mouse contains Lmbr1 surrounded by the orthologues of C7orf3 and C7orf4 and is ≈800 kb from Shh. No other genes are known to lie between Shh and the PPD critical region, suggesting that this is a gene poor region. The arrows indicate the transcriptional orientation. The structure of the 17 exon containing LMBR1 gene is depicted at the bottom and shows the relative position of each mutation. HPAP is the human placental alkaline phosphatase reporter gene contained within the transgene construct that resides at the Ssq insertion site. The double-headed arrows indicate the gene regions responsible for the opposing acheiropodia and PPD (Ssq) phenotypes. It is predicted that mutations between exons 5 and 6 release normal repression of the ectopic anterior expression pattern, whereas deletions around exon 4 inactivate limb-specific expression.
Similar articles
-
A novel candidate gene for mouse and human preaxial polydactyly with altered expression in limbs of Hemimelic extra-toes mutant mice.Genomics. 2000 Jul 1;67(1):19-27. doi: 10.1006/geno.2000.6225. Genomics. 2000. PMID: 10945466
-
Elimination of a long-range cis-regulatory module causes complete loss of limb-specific Shh expression and truncation of the mouse limb.Development. 2005 Feb;132(4):797-803. doi: 10.1242/dev.01613. Development. 2005. PMID: 15677727
-
Single base pair change in the long-range Sonic hedgehog limb-specific enhancer is a genetic basis for preaxial polydactyly.Dev Dyn. 2005 Feb;232(2):345-8. doi: 10.1002/dvdy.20254. Dev Dyn. 2005. PMID: 15637698
-
[Progress on polydactyly character of vertebrate].Yi Chuan. 2004 May;26(3):387-93. Yi Chuan. 2004. PMID: 15640026 Review. Chinese.
-
Sonic hedgehog: restricted expression and limb dysmorphologies.J Anat. 2003 Jan;202(1):13-20. doi: 10.1046/j.1469-7580.2003.00148.x. J Anat. 2003. PMID: 12587915 Free PMC article. Review.
Cited by
-
Extensive promoter-centered chromatin interactions provide a topological basis for transcription regulation.Cell. 2012 Jan 20;148(1-2):84-98. doi: 10.1016/j.cell.2011.12.014. Cell. 2012. PMID: 22265404 Free PMC article.
-
A new locus for split hand/foot malformation with long bone deficiency (SHFLD) at 2q14.2 identified from a chromosome translocation.Hum Genet. 2007 Sep;122(2):191-9. doi: 10.1007/s00439-007-0390-7. Epub 2007 Jun 14. Hum Genet. 2007. PMID: 17569090
-
Why study human limb malformations?J Anat. 2003 Jan;202(1):27-35. doi: 10.1046/j.1469-7580.2003.00130.x. J Anat. 2003. PMID: 12587917 Free PMC article.
-
A high-resolution map of the three-dimensional chromatin interactome in human cells.Nature. 2013 Nov 14;503(7475):290-4. doi: 10.1038/nature12644. Epub 2013 Oct 20. Nature. 2013. PMID: 24141950 Free PMC article.
-
Conserved noncoding elements follow power-law-like distributions in several genomes as a result of genome dynamics.PLoS One. 2014 May 2;9(5):e95437. doi: 10.1371/journal.pone.0095437. eCollection 2014. PLoS One. 2014. PMID: 24787386 Free PMC article.
References
-
- Heutink P, Zguricas J, van Oosterhout L, Breedveld G J, Testers L, Sandkuijl L A, Snijders P J, Weissenbach J, Lindhout D, Hovius S E, et al. Nat Genet. 1994;6:287–292. - PubMed
-
- Hing A V, Helms C, Slaugh R, Burgess A, Wang J C, Herman T, Dowton S B, Donis-Keller H. Am J Med Genet. 1995;58:128–135. - PubMed
-
- Heus H C, Hing A, van Baren M J, Joosse M, Breedveld G J, Wang J C, Burgess A, Donnis-Keller H, Berglund C, Zguricas J, et al. Genomics. 1999;57:342–351. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
