

Brain abnormalities, defective meiotic chromosome synapsis and female subfertility in HSF2 null mice
- PMID: 12032072
- PMCID: PMC125382
- DOI: 10.1093/emboj/21.11.2591
Brain abnormalities, defective meiotic chromosome synapsis and female subfertility in HSF2 null mice
Abstract
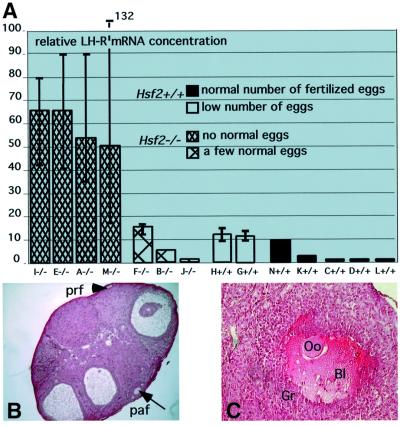
Heat shock factor 2, one of the four vertebrate HSFs, transcriptional regulators of heat shock gene expression, is active during embryogenesis and spermatogenesis, with unknown functions and targets. By disrupting the Hsf2 gene, we show that, although the lack of HSF2 is not embryonic lethal, Hsf2(-/-) mice suffer from brain abnormalities, and meiotic and gameto genesis defects in both genders. The disturbances in brain are characterized by the enlargement of lateral and third ventricles and the reduction of hippocampus and striatum, in correlation with HSF2 expression in proliferative cells of the neuroepithelium and in some ependymal cells in adults. Many developing spermatocytes are eliminated via apoptosis in a stage-specific manner in Hsf2(-/-) males, and pachytene spermatocytes also display structural defects in the synaptonemal complexes between homologous chromosomes. Hsf2(-/-) females suffer from multiple fertility defects: the production of abnormal eggs, the reduction in ovarian follicle number and the presence of hemorrhagic cystic follicles are consistent with meiotic defects. Hsf2(-/-) females also display hormone response defects, that can be rescued by superovulation treatment, and exhibit abnormal rates of luteinizing hormone receptor mRNAs.
Figures



Similar articles
-
Targeted disruption of the heat shock transcription factor (hsf)-2 gene results in increased embryonic lethality, neuronal defects, and reduced spermatogenesis.Genesis. 2003 May;36(1):48-61. doi: 10.1002/gene.10200. Genesis. 2003. PMID: 12748967
-
Essential requirement for both hsf1 and hsf2 transcriptional activity in spermatogenesis and male fertility.Genesis. 2004 Feb;38(2):66-80. doi: 10.1002/gene.20005. Genesis. 2004. PMID: 14994269
-
Stage-specific expression and cellular localization of the heat shock factor 2 isoforms in the rat seminiferous epithelium.Exp Cell Res. 1998 Apr 10;240(1):16-27. doi: 10.1006/excr.1997.3926. Exp Cell Res. 1998. PMID: 9570917
-
The Role of Heat Shock Factors in Mammalian Spermatogenesis.Adv Anat Embryol Cell Biol. 2017;222:45-65. doi: 10.1007/978-3-319-51409-3_3. Adv Anat Embryol Cell Biol. 2017. PMID: 28389750 Review.
-
Heat Shock Transcription Factor 2 Is Significantly Involved in Neurodegenerative Diseases, Inflammatory Bowel Disease, Cancer, Male Infertility, and Fetal Alcohol Spectrum Disorder: The Novel Mechanisms of Several Severe Diseases.Int J Mol Sci. 2022 Nov 9;23(22):13763. doi: 10.3390/ijms232213763. Int J Mol Sci. 2022. PMID: 36430241 Free PMC article. Review.
Cited by
-
Transcription and post-transcriptional regulation of spermatogenesis.Philos Trans R Soc Lond B Biol Sci. 2010 May 27;365(1546):1637-51. doi: 10.1098/rstb.2009.0196. Philos Trans R Soc Lond B Biol Sci. 2010. PMID: 20403875 Free PMC article. Review.
-
Association and regulation of heat shock transcription factor 4b with both extracellular signal-regulated kinase mitogen-activated protein kinase and dual-specificity tyrosine phosphatase DUSP26.Mol Cell Biol. 2006 Apr;26(8):3282-94. doi: 10.1128/MCB.26.8.3282-3294.2006. Mol Cell Biol. 2006. PMID: 16581800 Free PMC article.
-
Cell Cycle Regulation by Heat Shock Transcription Factors.Cells. 2022 Jan 8;11(2):203. doi: 10.3390/cells11020203. Cells. 2022. PMID: 35053319 Free PMC article. Review.
-
Advances of Heat Shock Family in Ulcerative Colitis.Front Pharmacol. 2022 May 12;13:869930. doi: 10.3389/fphar.2022.869930. eCollection 2022. Front Pharmacol. 2022. PMID: 35645809 Free PMC article. Review.
-
Meta-analysis of genome-wide association studies identifies novel loci that influence cupping and the glaucomatous process.Nat Commun. 2014 Sep 22;5:4883. doi: 10.1038/ncomms5883. Nat Commun. 2014. PMID: 25241763 Free PMC article.
References
-
- Alastalo T.P., Lonnstrom,M., Leppa,S., Kaarniranta,K., Pelto-Huikko,M., Sistonen,L. and Parvinen,M. (1998) Stage-specific expression and cellular localization of the heat shock factor 2 isoforms in the rat seminiferous epithelium. Exp. Cell Res., 240, 16–27. - PubMed
-
- Baudat F., Manova,K., Pui Yuen,J., Jasin,M. and Keeney,S. (2000) Chromosone synapsis defects and sexually dimorphic meiotic progression in mice lacking Spo11. Mol. Cell, 6, 989–998. - PubMed
-
- Christians E., Davis,A.A., Thomas,S.D. and Benjamin,I.J. (2000) Maternal effect of Hsf1 on reproductive success. Nature, 407, 693–694. - PubMed
-
- Clermont Y. (1972) Kinetics of spermatogenesis in mammals: seminiferous epithelium cycle and spermatogonia renewal. Physiol. Rev., 52, 198–236. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
