

Connected gene neighborhoods in prokaryotic genomes
- PMID: 12000841
- PMCID: PMC115289
- DOI: 10.1093/nar/30.10.2212
Connected gene neighborhoods in prokaryotic genomes
Abstract
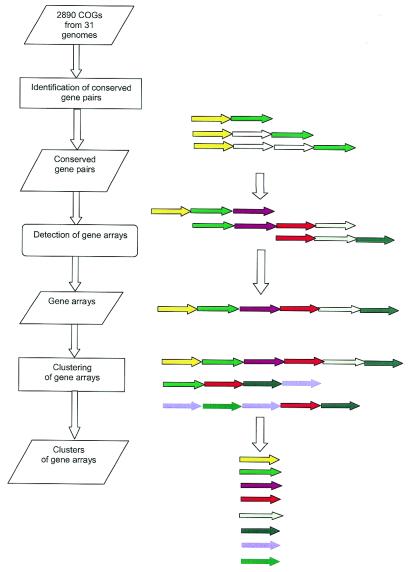
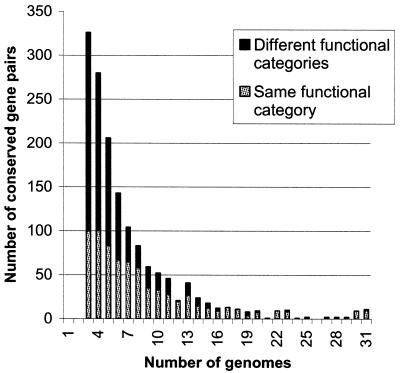
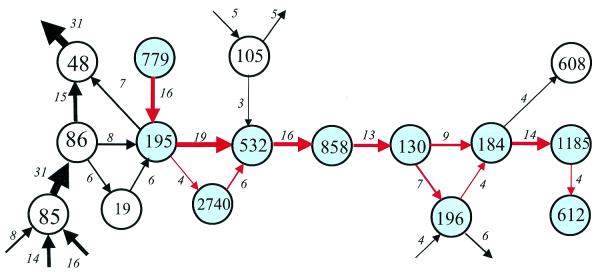
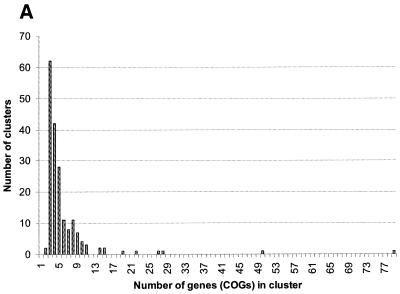
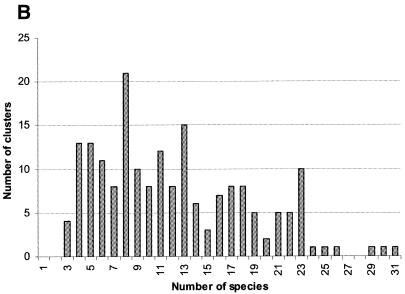
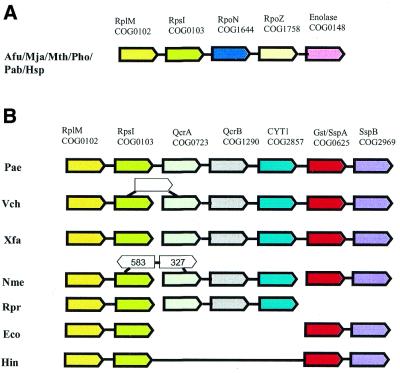
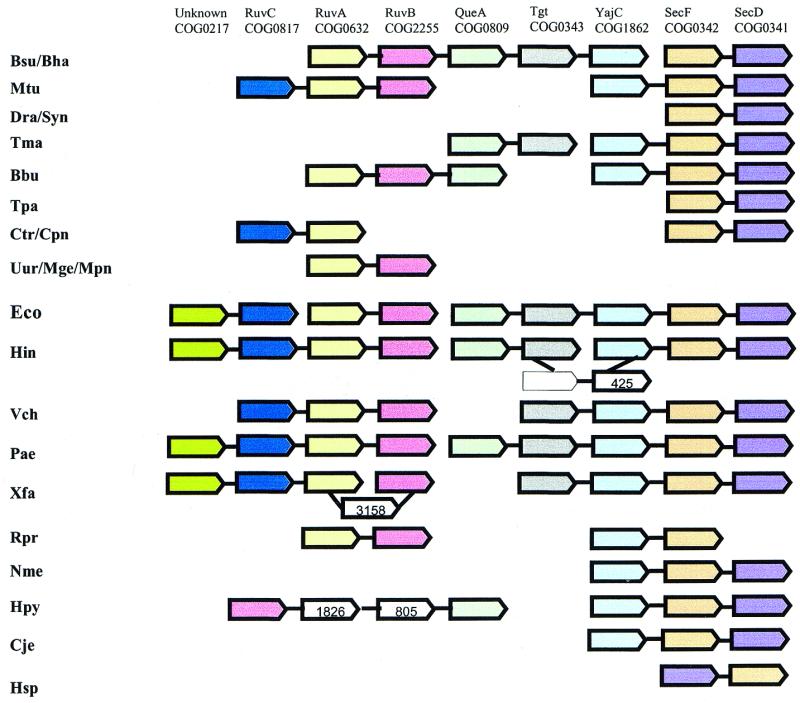
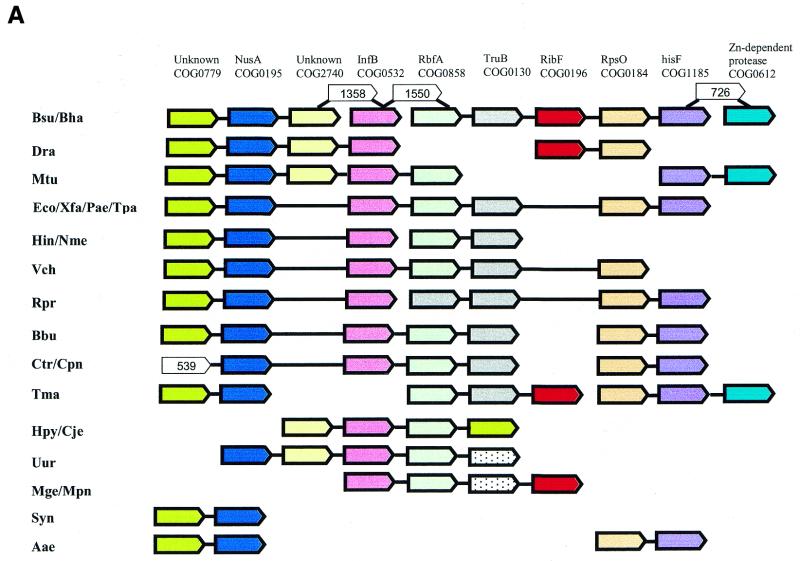
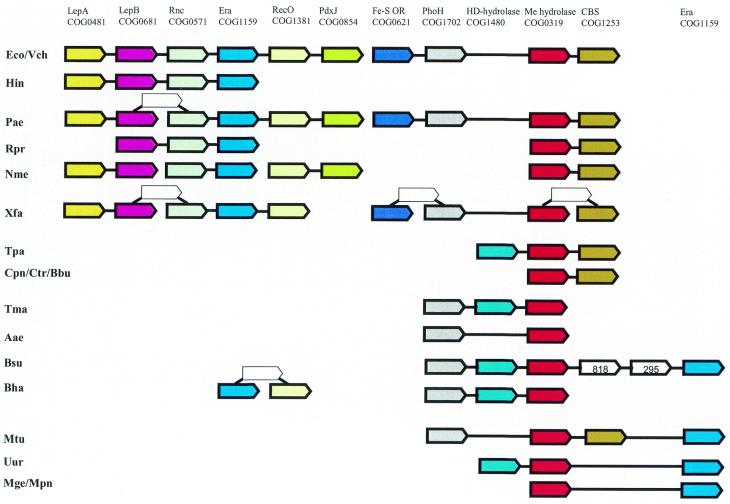
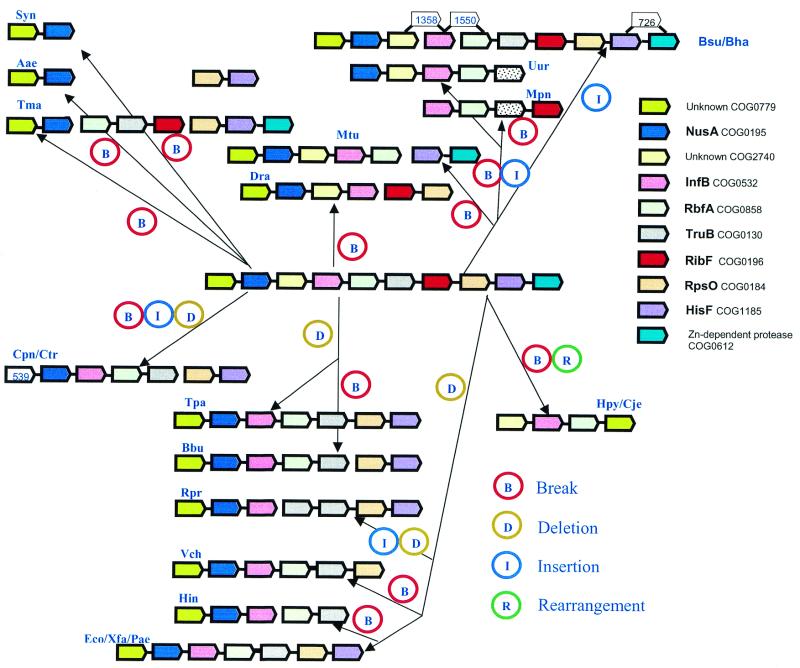
A computational method was developed for delineating connected gene neighborhoods in bacterial and archaeal genomes. These gene neighborhoods are not typically present, in their entirety, in any single genome, but are held together by overlapping, partially conserved gene arrays. The procedure was applied to comparing the orders of orthologous genes, which were extracted from the database of Clusters of Orthologous Groups of proteins (COGs), in 31 prokaryotic genomes and resulted in the identification of 188 clusters of gene arrays, which included 1001 of 2890 COGs. These clusters were projected onto actual genomes to produce extended neighborhoods including additional genes, which are adjacent to the genes from the clusters and are transcribed in the same direction, which resulted in a total of 2387 COGs being included in the neighborhoods. Most of the neighborhoods consist predominantly of genes united by a coherent functional theme, but also include a minority of genes without an obvious functional connection to the main theme. We hypothesize that although some of the latter genes might have unsuspected roles, others are maintained within gene arrays because of the advantage of expression at a level that is typical of the given neighborhood. We designate this phenomenon 'genomic hitchhiking'. The largest neighborhood includes 79 genes (COGs) and consists of overlapping, rearranged ribosomal protein superoperons; apparent genome hitchhiking is particularly typical of this neighborhood and other neighborhoods that consist of genes coding for translation machinery components. Several neighborhoods involve previously undetected connections between genes, allowing new functional predictions. Gene neighborhoods appear to evolve via complex rearrangement, with different combinations of genes from a neighborhood fixed in different lineages.
Figures

Similar articles
-
Clusters of orthologous genes for 41 archaeal genomes and implications for evolutionary genomics of archaea.Biol Direct. 2007 Nov 27;2:33. doi: 10.1186/1745-6150-2-33. Biol Direct. 2007. PMID: 18042280 Free PMC article.
-
Genome alignment, evolution of prokaryotic genome organization, and prediction of gene function using genomic context.Genome Res. 2001 Mar;11(3):356-72. doi: 10.1101/gr.gr-1619r. Genome Res. 2001. PMID: 11230160
-
Genome trees constructed using five different approaches suggest new major bacterial clades.BMC Evol Biol. 2001 Oct 20;1:8. doi: 10.1186/1471-2148-1-8. BMC Evol Biol. 2001. PMID: 11734060 Free PMC article.
-
A genomic perspective on protein families.Science. 1997 Oct 24;278(5338):631-7. doi: 10.1126/science.278.5338.631. Science. 1997. PMID: 9381173 Review.
-
Comparative Genomics for Prokaryotes.Methods Mol Biol. 2018;1704:55-78. doi: 10.1007/978-1-4939-7463-4_3. Methods Mol Biol. 2018. PMID: 29277863 Review.
Cited by
-
A non-random walk through the genome.Genome Biol. 2005;6(4):214. doi: 10.1186/gb-2005-6-4-214. Epub 2005 Mar 31. Genome Biol. 2005. PMID: 15833129 Free PMC article. Review.
-
Operon formation is driven by co-regulation and not by horizontal gene transfer.Genome Res. 2005 Jun;15(6):809-19. doi: 10.1101/gr.3368805. Genome Res. 2005. PMID: 15930492 Free PMC article.
-
Clusters of orthologous genes for 41 archaeal genomes and implications for evolutionary genomics of archaea.Biol Direct. 2007 Nov 27;2:33. doi: 10.1186/1745-6150-2-33. Biol Direct. 2007. PMID: 18042280 Free PMC article.
-
Resistance-Guided Mining of Bacterial Genotoxins Defines a Family of DNA Glycosylases.mBio. 2022 Apr 26;13(2):e0329721. doi: 10.1128/mbio.03297-21. Epub 2022 Mar 21. mBio. 2022. PMID: 35311535 Free PMC article.
-
Computational approaches to protein-protein interaction.J Struct Funct Genomics. 2003;4(4):245-55. doi: 10.1023/b:jsfg.0000016143.91973.1c. J Struct Funct Genomics. 2003. PMID: 15185965
References
-
- Jacob F., Perrin,D., Sanchez,C. and Monod,J. (1960) L’Operon: groupe de genes a expression coordonee par un operateur. C. R. Acad. Sci., 250, 1727–1729. - PubMed
-
- Jacob F. and Monod,J. (1961) Genetic regulatory mechanisms in the synthesis of proteins. J. Mol. Biol., 3, 318–356. - PubMed
-
- Miller J.H. and Reznikoff,W.S.E. (1978) The Operon. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
-
- Mushegian A.R. and Koonin,E.V. (1996) Gene order is not conserved in bacterial evolution. Trends Genet., 12, 289–290. - PubMed
-
- Dandekar T., Snel,B., Huynen,M. and Bork,P. (1998) Conservation of gene order: a fingerprint of proteins that physically interact. Trends Biochem. Sci., 23, 324–328. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
