

Identification of myogenic-endothelial progenitor cells in the interstitial spaces of skeletal muscle
- PMID: 11994315
- PMCID: PMC2173851
- DOI: 10.1083/jcb.200112106
Identification of myogenic-endothelial progenitor cells in the interstitial spaces of skeletal muscle
Abstract
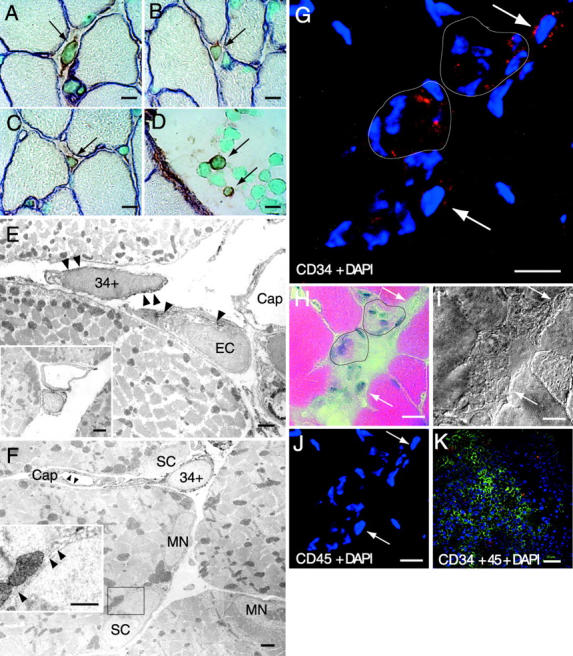
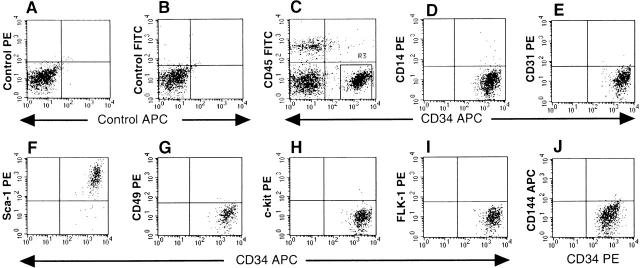
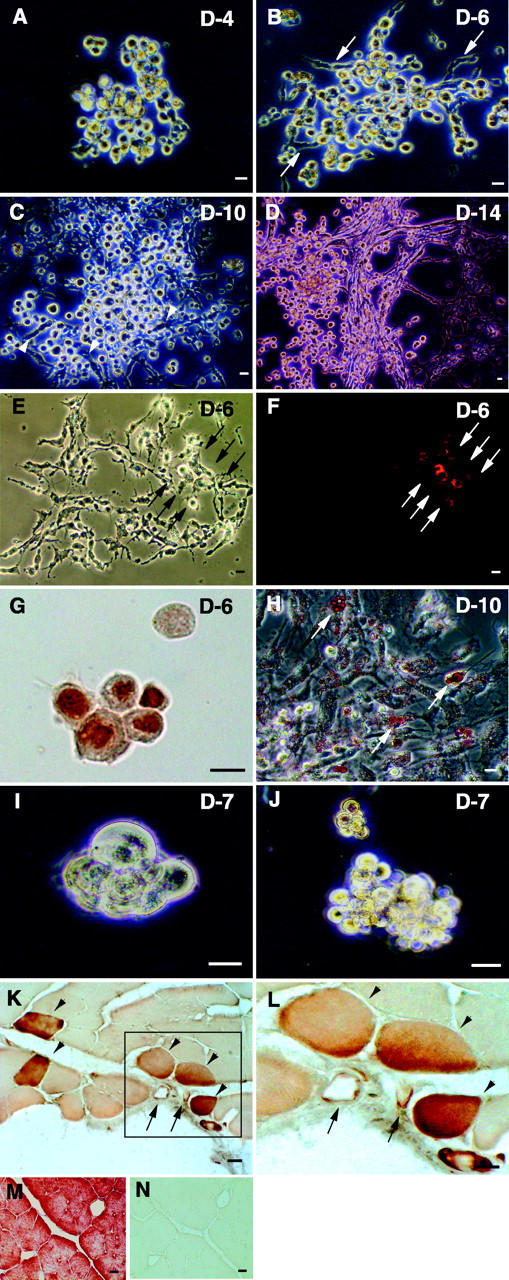
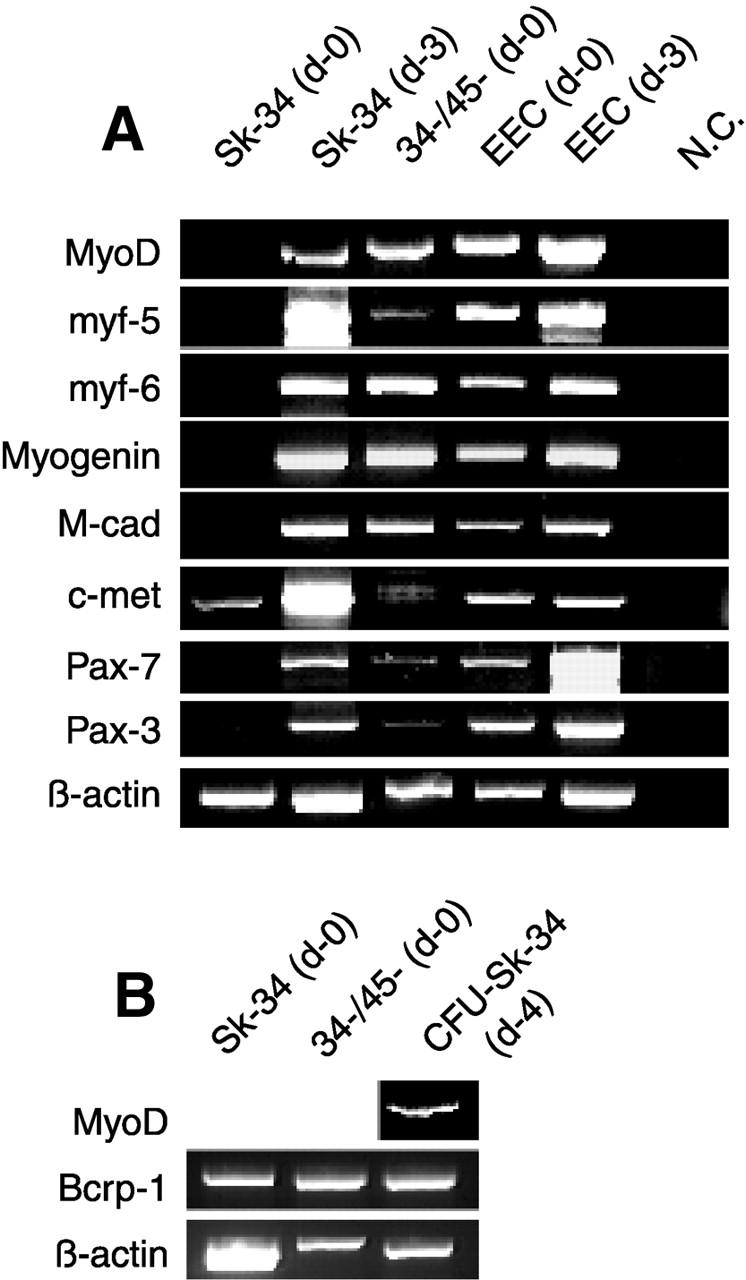
Putative myogenic and endothelial (myo-endothelial) cell progenitors were identified in the interstitial spaces of murine skeletal muscle by immunohistochemistry and immunoelectron microscopy using CD34 antigen. Enzymatically isolated cells were characterized by fluorescence-activated cell sorting on the basis of cell surface antigen expression, and were sorted as a CD34+ and CD45- fraction. Cells in this fraction were approximately 94% positive for Sca-1, and mostly negative (<3% positive) for CD14, 31, 49, 144, c-kit, and FLK-1. The CD34+/45- cells formed colonies in clonal cell cultures and colony-forming units displayed the potential to differentiate into adipocytes, endothelial, and myogenic cells. The CD34+/45- cells fully differentiated into vascular endothelial cells and skeletal muscle fibers in vivo after transplantation. Immediately after sorting, CD34+/45- cells expressed only c-met mRNA, and did not express any other myogenic cell-related markers such as MyoD, myf-5, myf-6, myogenin, M-cadherin, Pax-3, and Pax-7. However, after 3 d of culture, these cells expressed mRNA for all myogenic markers. CD34+/45- cells were distinct from satellite cells, as they expressed Bcrp1/ABCG2 gene mRNA (Zhou et al., 2001). These findings suggest that myo-endothelial progenitors reside in the interstitial spaces of mammalian skeletal muscles, and that they can potentially contribute to postnatal skeletal muscle growth.
Figures
Similar articles
-
Growth and differentiation potential of main- and side-population cells derived from murine skeletal muscle.Exp Cell Res. 2003 Nov 15;291(1):83-90. doi: 10.1016/s0014-4827(03)00376-8. Exp Cell Res. 2003. PMID: 14597410
-
New fiber formation in the interstitial spaces of rat skeletal muscle during postnatal growth.J Histochem Cytochem. 2002 Aug;50(8):1097-111. doi: 10.1177/002215540205000812. J Histochem Cytochem. 2002. PMID: 12133913
-
The role of CD34 expression and cellular fusion in the regeneration capacity of myogenic progenitor cells.J Cell Sci. 2002 Nov 15;115(Pt 22):4361-74. doi: 10.1242/jcs.00110. J Cell Sci. 2002. PMID: 12376567
-
Plasticity and physiological role of stem cells derived from skeletal muscle interstitium: contribution to muscle fiber hyperplasia and therapeutic use.Curr Pharm Des. 2010;16(8):956-67. doi: 10.2174/138161210790883408. Curr Pharm Des. 2010. PMID: 20041822 Review.
-
Concise review: evidence for CD34 as a common marker for diverse progenitors.Stem Cells. 2014 Jun;32(6):1380-9. doi: 10.1002/stem.1661. Stem Cells. 2014. PMID: 24497003 Free PMC article. Review.
Cited by
-
Regenerative Translation of Human Blood-Vessel-Derived MSC Precursors.Stem Cells Int. 2015;2015:375187. doi: 10.1155/2015/375187. Epub 2015 Jul 26. Stem Cells Int. 2015. PMID: 26273304 Free PMC article. Review.
-
Endothelial progenitor cells for postnatal vasculogenesis.Clin Exp Nephrol. 2007 Mar;11(1):18-25. doi: 10.1007/s10157-006-0448-1. Epub 2007 Mar 28. Clin Exp Nephrol. 2007. PMID: 17384994 Review.
-
Primary rat muscle progenitor cells have decreased proliferation and myotube formation during passages.Cell Prolif. 2004 Aug;37(4):267-77. doi: 10.1111/j.1365-2184.2004.00311.x. Cell Prolif. 2004. PMID: 15245563 Free PMC article.
-
Denervated muscle extract promotes recovery of muscle atrophy through activation of satellite cells. An experimental study.J Sport Health Sci. 2019 Jan;8(1):23-31. doi: 10.1016/j.jshs.2017.05.007. Epub 2017 May 26. J Sport Health Sci. 2019. PMID: 30719380 Free PMC article.
-
Direct isolation, culture and transplant of mouse skeletal muscle derived endothelial cells with angiogenic potential.PLoS One. 2008 Mar 12;3(3):e0001753. doi: 10.1371/journal.pone.0001753. PLoS One. 2008. PMID: 18335025 Free PMC article.
References
-
- Asahara, T., T. Murohara, A. Sullivan, M. Silver, R. van der Zee, T. Li, B. Witzenbichler, G. Schatteman, and J.M. Isner. 1997. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 275:964–967. - PubMed
-
- Birchmeier, C., and E. Gherardi. 1998. Developmental roles of HGF/SF and its receptor, the c-Met tyrosine kinase. Trends Cell Biol. 8:404–410. - PubMed
-
- Bischoff, R. 1986. Proliferation of muscle satellite cells on intact myofibers in culture. Dev. Biol. 115:129–139. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
