

doi: 10.1073/pnas.102483199.
Epub 2002 Apr 30.
The OhrR repressor senses organic hydroperoxides by reversible formation of a cysteine-sulfenic acid derivative
Affiliations
- PMID: 11983871
- PMCID: PMC124464
- DOI: 10.1073/pnas.102483199
Item in Clipboard
The OhrR repressor senses organic hydroperoxides by reversible formation of a cysteine-sulfenic acid derivative
Proc Natl Acad Sci U S A.
.
Abstract
Reactive oxygen species induce the expression of detoxification and repair genes critical for life in an aerobic environment. Bacterial factors that sense reactive oxygen species use either thiol-disulfide exchange reactions (OxyR, RsrA) or redox labile 2Fe-2S clusters (SoxR). We demonstrate that the reduced form of Bacillus subtilis OhrR binds cooperatively to two adjacent inverted repeat sequences in the ohrA control region and thereby represses transcription. In the presence of organic hydroperoxides, OhrR is inactivated by the reversible oxidation of a single conserved cysteine residue to the corresponding cysteine-sulfenic acid, and perhaps to higher oxidation states.
Figures
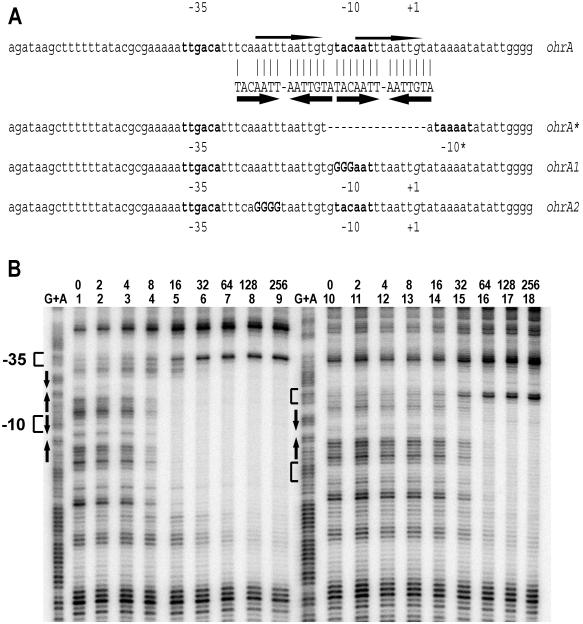
Binding of OhrR to the ohrA and ohrA* promoters. (A) The ohrA promoter and variants used in this study. The wild-type promoter (ohrA) contains two inverted repeats shown by thick arrows. Bases matching the perfect inverted repeat are identified by a vertical line. The thin arrows indicate the 11-bp direct repeats. The −10 and −35 regions are shown in bold letters, and the transcription start site (+1 position) is in italics. ohrA* is a previously described 15-bp deletion in the promoter region (10) as indicated with a dashed line. The ohrA1 mutation (uppercase bold letters) destroys the perfect inverted repeat (but not the direct repeat). The ohrA2 mutation (uppercase bold letters) destroys the direct repeat as well as the imperfect inverted repeat. −10* is a new −10 region in the ohrA* promoter. (B) DNase I footprinting analysis of OhrR binding to the ohrA (lanes 1–9) and ohrA* (lanes 10–18) promoters. Both promoter regions (2 nM DNA) were labeled on the top strand, and the amount of OhrR (nM) is shown above the lane number. Note that at high concentrations the protein binding site extends in the downstream direction. DTT (1 mM) was present in all binding reactions. The −10, −35, and +1 positions are shown next to the G+A ladders. The inverted repeats are shown with arrows.
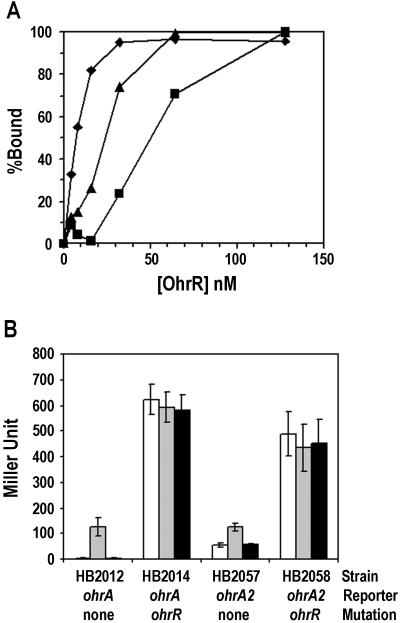
Binding of OhrR to mutant ohrA operators in vitro and in vivo. (A) The binding of OhrR (% DNA bound) to ohrA (⧫), ohrA1 (▪), and ohrA2 (▴) promoter DNA as determined from DNaseI footprinting data (promoter DNA at 1 nM). (B) In vivo expression from reporter fusions for the ohrA and ohrA2 promoters. Log phase cultures of each strain were induced with 100 μM CHP (gray bar) or 100 μM H2O2 (black bar) for 15 min at 37°C with aeration before β-galactosidase assay. Empty bars are nontreated cultures. Results are means ± SD from three independent experiments performed in duplicate. Note that ohrA1 promoter activity, which is not shown here, gave less than 1 Miller unit.
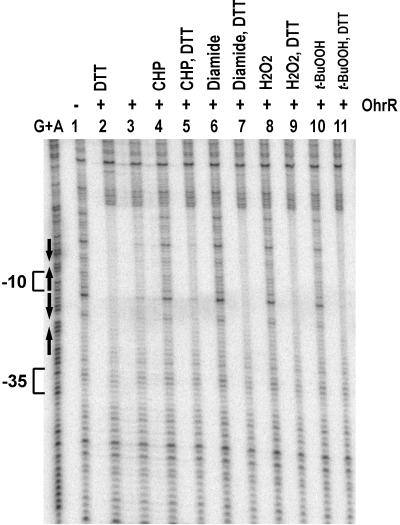
Effects of oxidants on binding of OhrR to the ohrA promoter. The fragment (0.1 nM DNA) used in this experiment was the same as in Fig. 2 except that AccI digestion was used to obtain the fragment for bottom-strand labeling using α-[32P]dATP. Either 1 mM CHP, 1 mM tert-butyl hydroperoxide (t-BuOOH), 1 mM H2O2, or 10 mM diamide was added to the binding reaction and incubated for 10 min at room temperature. When indicated, 100 mM DTT was then added into the reactions and incubation continued at room temperature for 30 min before DNaseI treatment. Lane 1 contained no protein; lanes 2–12 contained 2 nM OhrR.
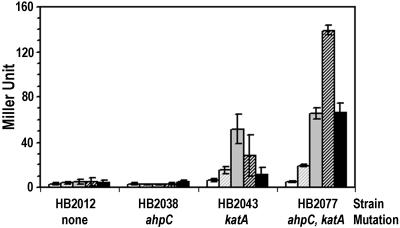
Induction of ohrA by H2O2. β-Galactosidase activity of strains containing the SPβc2Δ2∷Tn917∷φ(ohrA-cat-lacZ) reporter fusion were determined in wild type (HB2012), an ahpC∷Tn10 mutant (HB2038), a catalase mutant strain [HB2043; CU1065 katA∷pJH101 (32)], and the double mutant (HB2077). Samples were taken 15 min after treatment with H2O2 to a final concentration of (left to right) 0, 100, 200, 400, or 800 μM.
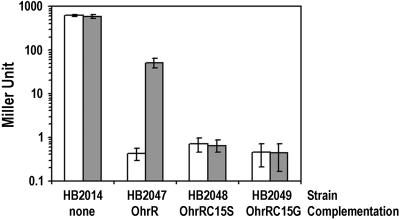
Essential role for OhrR Cys-15 in peroxide sensing. β-Galactosidase activity of strains containing ohrA-cat lacZ, ohrR∷kan complemented with OhrR, OhrR C15S, or OhrR C15G. Log-phase cultures of each strain were induced with 100 μM CHP for 15 min at 37°C with aeration before β-galactosidase assay. The empty bars are nontreated cultures, and the filled bars are CHP-treated cultures. Results are means ± SD from three independent experiments performed in duplicate.
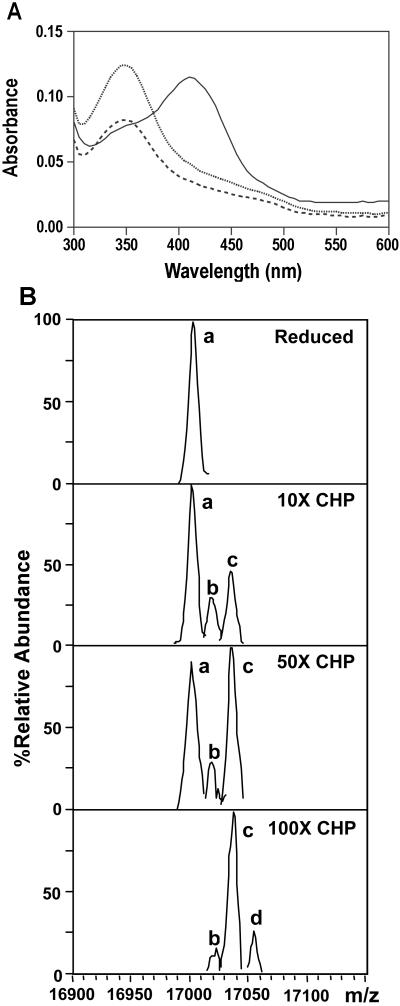
Sulfenic acid formation at C15 upon oxidative stress. (A) NBD-Cl modification of purified OhrR. Reduced OhrR was treated with CHP for 30 s (dotted line) and 10 min (dashed line). The thiol group and sulfenic acid were trapped by addition of NBD-Cl as described (22). The absorbance at 300–600 mm was scanned with a microtiter plate reader. The reaction without OhrR was used as blank. Solid line is reduced OhrR reacted with NBD-Cl. (B) Detection of OhrR oxidation by electrospray ionization–MS. Protein was incubated with CHP and immediately (within 30 s) infused into a Bruker Esquire electrospray ionization–MS instrument for analysis. The deconvoluted spectra were derived as described in Materials and Methods and peak heights were normalized to the most abundant species (set to 100%). Peak a corresponds in mass to the fully reduced protein, and peaks b, c, and d indicate the incorporation of 1, 2, or 3 oxygen atoms, respectively.
Similar articles
-
Oxidant-dependent switching between reversible and sacrificial oxidation pathways for Bacillus subtilis OhrR.Mol Microbiol. 2008 May;68(4):978-86. doi: 10.1111/j.1365-2958.2008.06200.x. Epub 2008 Mar 19. Mol Microbiol. 2008. PMID: 18363800
-
OhrR, a transcription repressor that senses and responds to changes in organic peroxide levels in Xanthomonas campestris pv. phaseoli.Mol Microbiol. 2002 Sep;45(6):1647-54. doi: 10.1046/j.1365-2958.2002.03116.x. Mol Microbiol. 2002. PMID: 12354231
-
A complex thiolate switch regulates the Bacillus subtilis organic peroxide sensor OhrR.Proc Natl Acad Sci U S A. 2007 May 22;104(21):8743-8. doi: 10.1073/pnas.0702081104. Epub 2007 May 14. Proc Natl Acad Sci U S A. 2007. PMID: 17502599 Free PMC article.
-
PerR vs OhrR: selective peroxide sensing in Bacillus subtilis.Mol Biosyst. 2010 Feb;6(2):316-23. doi: 10.1039/b915042k. Epub 2009 Sep 18. Mol Biosyst. 2010. PMID: 20094649 Review.
-
Regulation of inducible peroxide stress responses.Mol Microbiol. 2002 Jul;45(1):9-15. doi: 10.1046/j.1365-2958.2002.03015.x. Mol Microbiol. 2002. PMID: 12100544 Review.
Cited by
-
Regulation of MntH by a dual Mn(II)- and Fe(II)-dependent transcriptional repressor (DR2539) in Deinococcus radiodurans.PLoS One. 2012;7(4):e35057. doi: 10.1371/journal.pone.0035057. Epub 2012 Apr 16. PLoS One. 2012. PMID: 22523570 Free PMC article.
-
Chemical biology approaches to study protein cysteine sulfenylation.Biopolymers. 2014 Feb;101(2):165-72. doi: 10.1002/bip.22255. Biopolymers. 2014. PMID: 23576224 Free PMC article. Review.
-
Cys303 in the histidine kinase PhoR is crucial for the phosphotransfer reaction in the PhoPR two-component system in Bacillus subtilis.J Bacteriol. 2007 Jan;189(2):410-21. doi: 10.1128/JB.01205-06. Epub 2006 Nov 3. J Bacteriol. 2007. PMID: 17085571 Free PMC article.
-
Differential gene expression in response to hydrogen peroxide and the putative PerR regulon of Synechocystis sp. strain PCC 6803.J Bacteriol. 2004 Jun;186(11):3331-45. doi: 10.1128/JB.186.11.3331-3345.2004. J Bacteriol. 2004. PMID: 15150218 Free PMC article.
-
Measurement of protein sulfenic acid content.Curr Protoc Toxicol. 2008 Nov;Chapter 17:Unit17.2. doi: 10.1002/0471140856.tx1702s38. Curr Protoc Toxicol. 2008. PMID: 20963754 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
