

RNA-protein interactions that regulate pre-mRNA splicing
- PMID: 11868989
- PMCID: PMC5977533
RNA-protein interactions that regulate pre-mRNA splicing
Abstract
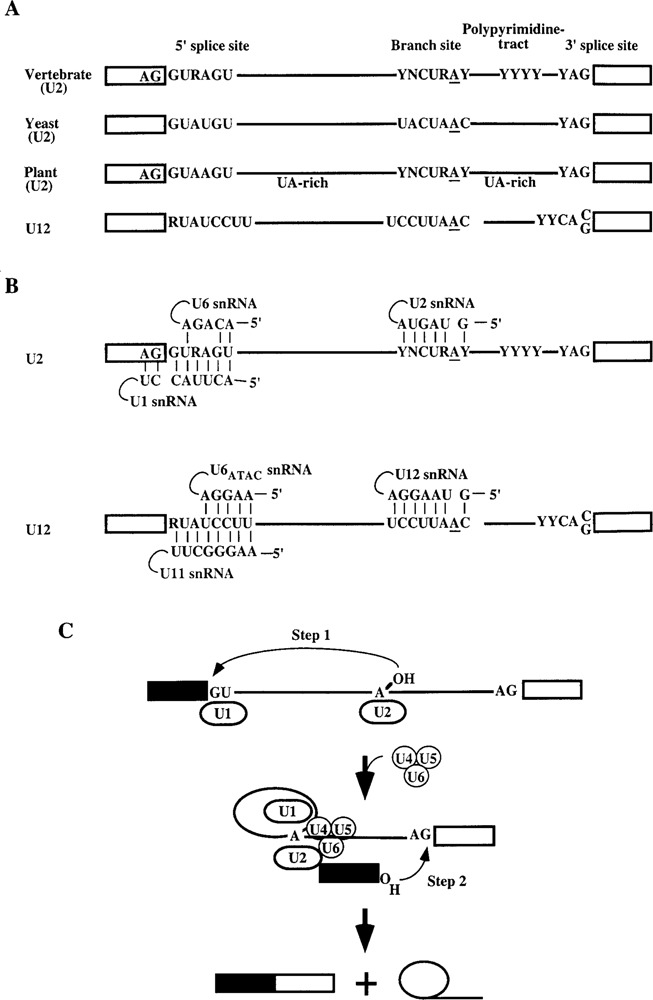
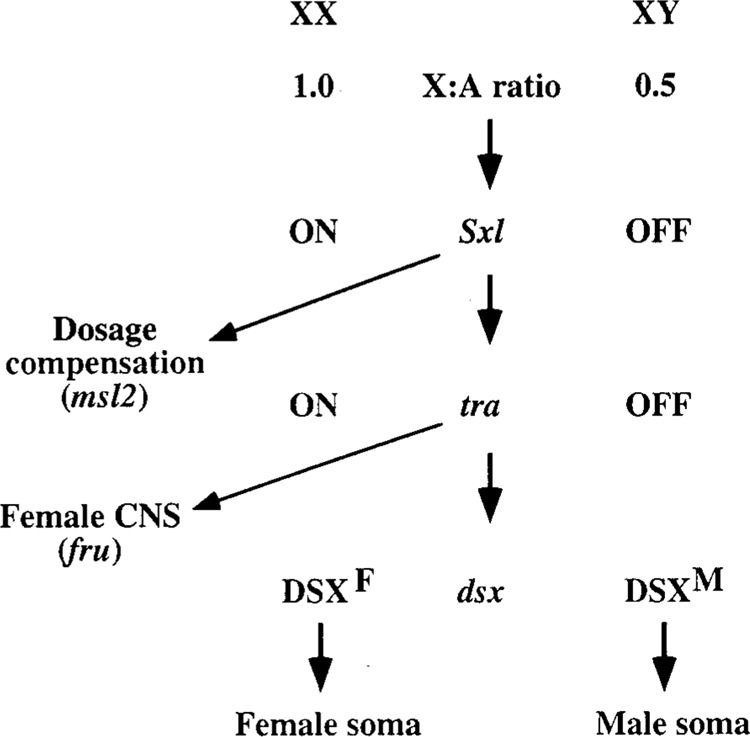
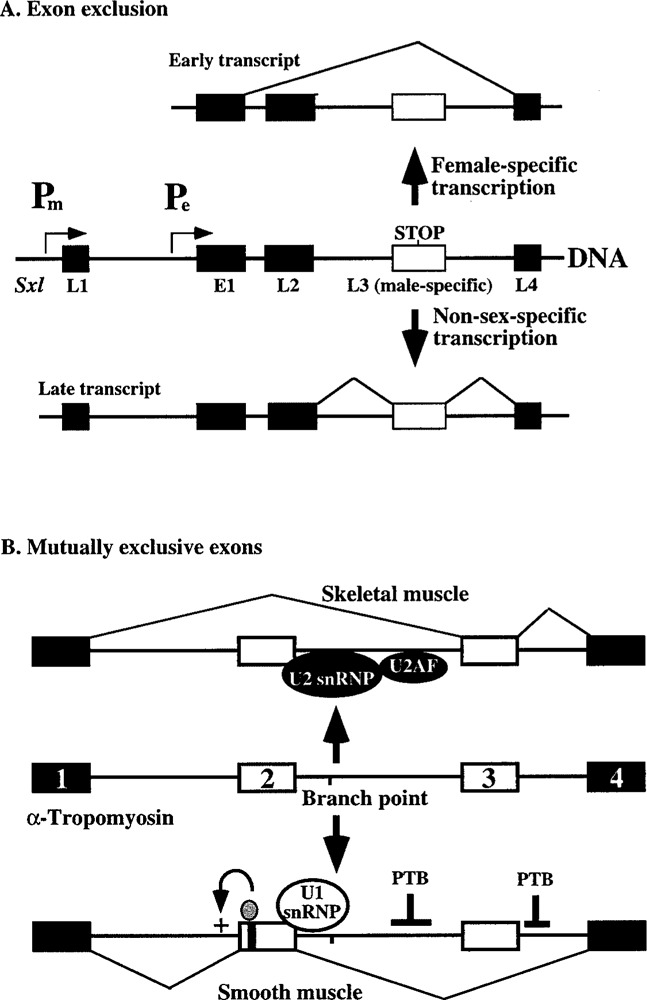
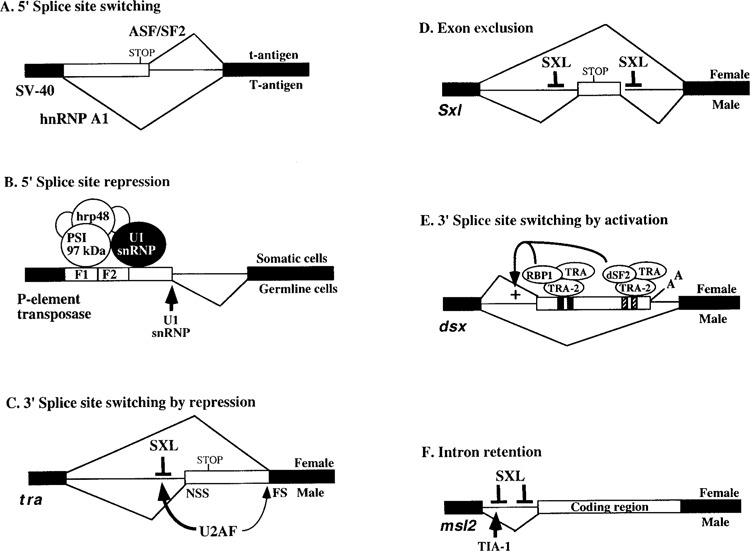
Splicing of nuclear precursor messenger RNAs is an important and ubiquitous type of gene regulation in metazoans. Splicing joins the coding sequences called exons by removing the intervening noncoding sequences, introns, from primary transcripts. Alternative splicing generates an enormous repertoire of functional diversity by producing multiple RNAs and proteins from a single gene. In fact, recent genome sequences from several organisms suggest that splicing regulation is likely to provide an important source of functional diversity in more complex organisms. Because splice sites are short sequences at the ends of introns, the functional splice sites have to be distinguished from an excessively large number of sequences in the primary transcripts that resemble a splice site. Furthermore, alternative splice sites have to be correctly chosen at appropriate times. Thus, selection of proper splice sites remains a daunting biological problem. This review focuses on a few examples in which the molecular and biochemical basis for splice site selection is better understood.
Figures
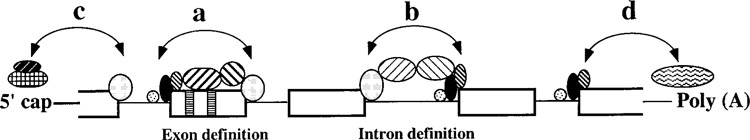
Similar articles
-
SR protein kinases: the splice of life.Biochem Cell Biol. 1999;77(4):293-8. Biochem Cell Biol. 1999. PMID: 10546892 Review.
-
Mechanisms of alternative pre-messenger RNA splicing.Annu Rev Biochem. 2003;72:291-336. doi: 10.1146/annurev.biochem.72.121801.161720. Epub 2003 Feb 27. Annu Rev Biochem. 2003. PMID: 12626338 Review.
-
P element temperature-specific transposition: a model for possible regulation of mobile elements activity by pre-mRNA secondary structure.Tsitol Genet. 2014 Nov-Dec;48(6):40-4. Tsitol Genet. 2014. PMID: 25536821
-
Cancer-Associated Perturbations in Alternative Pre-messenger RNA Splicing.Cancer Treat Res. 2013;158:41-94. doi: 10.1007/978-3-642-31659-3_3. Cancer Treat Res. 2013. PMID: 24222354 Review.
-
Pre-mRNA splicing: a complex picture in higher definition.Trends Biochem Sci. 2008 Jun;33(6):243-6. doi: 10.1016/j.tibs.2008.04.004. Epub 2008 May 9. Trends Biochem Sci. 2008. PMID: 18472266 Review.
Cited by
-
RPI-Pred: predicting ncRNA-protein interaction using sequence and structural information.Nucleic Acids Res. 2015 Feb 18;43(3):1370-9. doi: 10.1093/nar/gkv020. Epub 2015 Jan 21. Nucleic Acids Res. 2015. PMID: 25609700 Free PMC article.
-
Diverse Mechanisms of RNA Recombination.Mol Biol. 2005;39(4):529-542. doi: 10.1007/s11008-005-0069-x. Mol Biol. 2005. PMID: 32214466 Free PMC article.
-
The conserved RNA recognition motif 3 of U2 snRNA auxiliary factor (U2AF 65) is essential in vivo but dispensable for activity in vitro.RNA. 2004 Feb;10(2):240-53. doi: 10.1261/rna.5153204. RNA. 2004. PMID: 14730023 Free PMC article.
-
A novel method for the identification of conserved structural patterns in RNA: From small scale to high-throughput applications.Nucleic Acids Res. 2016 Oct 14;44(18):8600-8609. doi: 10.1093/nar/gkw750. Epub 2016 Aug 31. Nucleic Acids Res. 2016. PMID: 27580722 Free PMC article.
-
Prediction of RNA-binding proteins from primary sequence by a support vector machine approach.RNA. 2004 Mar;10(3):355-68. doi: 10.1261/rna.5890304. RNA. 2004. PMID: 14970381 Free PMC article.
References
-
- Adams M. D.; Celniker S. E.; Holt R. A.; et al. The genome sequence of Drosophila melanogaster . Science 287:2185–2195; 2000. - PubMed
-
- Bashaw G. J.; Baker B. S. The regulation of the Drosophila msi-2 gene reveals a function for Sex-lethal in translational control. Cell 89:789–798; 1997. - PubMed
-
- Berget S. M. Exon recognition in vertebrate splicing. J. Biol. Chem. 270:2411–2414; 1995. - PubMed
-
- Black D. L. Protein diversity from alternative splicing: A challenge for bioinformatics and post-genome biology. Cell 103:367–370; 2000. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources