

Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid
- PMID: 11842171
- PMCID: PMC148929
- DOI: 10.1104/pp.010320
Protection against heat stress-induced oxidative damage in Arabidopsis involves calcium, abscisic acid, ethylene, and salicylic acid
Abstract
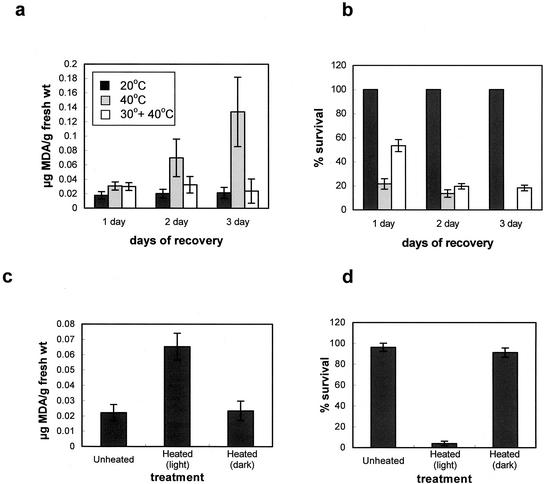
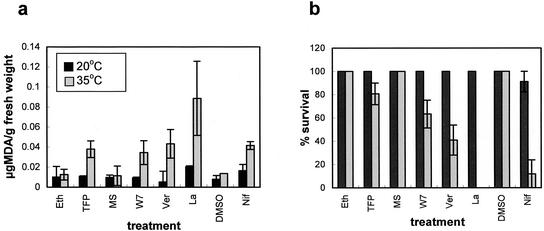
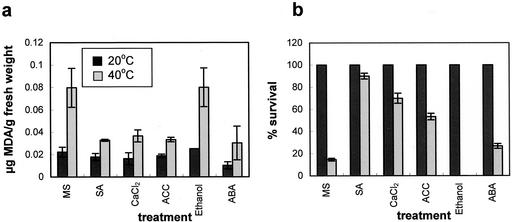
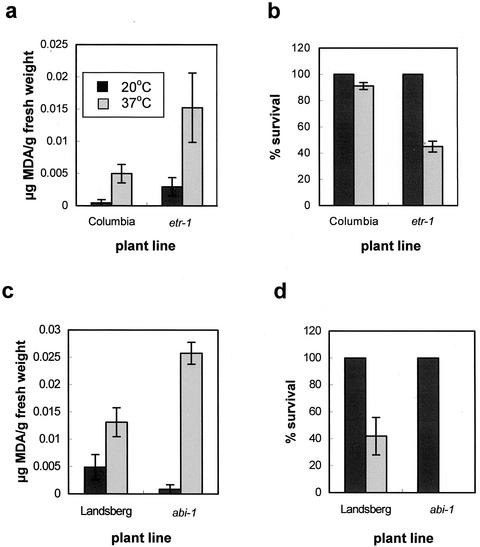
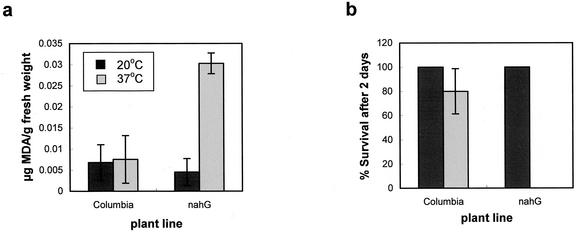
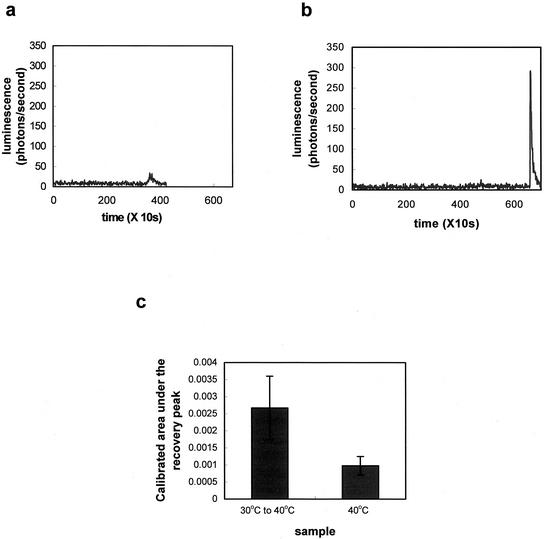
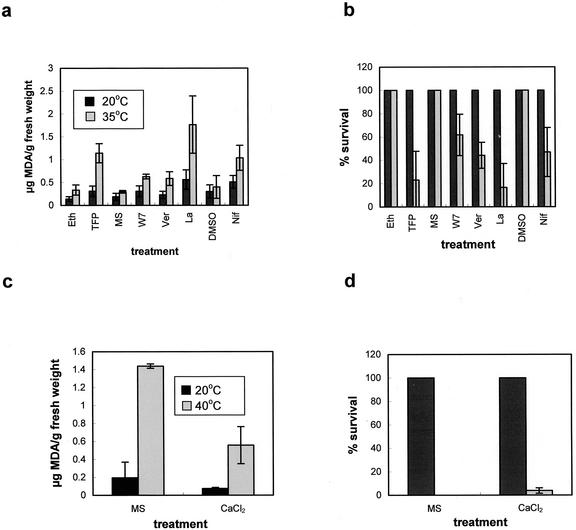
Plants, in common with all organisms, have evolved mechanisms to cope with the problems caused by high temperatures. We examined specifically the involvement of calcium, abscisic acid (ABA), ethylene, and salicylic acid (SA) in the protection against heat-induced oxidative damage in Arabidopsis. Heat caused increased thiobarbituric acid reactive substance levels (an indicator of oxidative damage to membranes) and reduced survival. Both effects required light and were reduced in plants that had acquired thermotolerance through a mild heat pretreatment. Calcium channel blockers and calmodulin inhibitors increased these effects of heating and added calcium reversed them, implying that protection against heat-induced oxidative damage in Arabidopsis requires calcium and calmodulin. Similar to calcium, SA, 1-aminocyclopropane-1-carboxylic acid (a precursor to ethylene), and ABA added to plants protected them from heat-induced oxidative damage. In addition, the ethylene-insensitive mutant etr-1, the ABA-insensitive mutant abi-1, and a transgenic line expressing nahG (consequently inhibited in SA production) showed increased susceptibility to heat. These data suggest that protection against heat-induced oxidative damage in Arabidopsis also involves ethylene, ABA, and SA. Real time measurements of cytosolic calcium levels during heating in Arabidopsis detected no increases in response to heat per se, but showed transient elevations in response to recovery from heating. The magnitude of these calcium peaks was greater in thermotolerant plants, implying that these calcium signals might play a role in mediating the effects of acquired thermotolerance. Calcium channel blockers and calmodulin inhibitors added solely during the recovery phase suggest that this role for calcium is in protecting against oxidative damage specifically during/after recovery.
Figures
Similar articles
-
Thermotolerance and antioxidant systems in Agrostis stolonifera: involvement of salicylic acid, abscisic acid, calcium, hydrogen peroxide, and ethylene.J Plant Physiol. 2004 Apr;161(4):405-13. doi: 10.1078/0176-1617-01239. J Plant Physiol. 2004. PMID: 15128028
-
Proline accumulation is inhibitory to Arabidopsis seedlings during heat stress.Plant Physiol. 2011 Aug;156(4):1921-33. doi: 10.1104/pp.111.175810. Epub 2011 Jun 13. Plant Physiol. 2011. PMID: 21670222 Free PMC article.
-
Heat stress phenotypes of Arabidopsis mutants implicate multiple signaling pathways in the acquisition of thermotolerance.Plant Physiol. 2005 Jun;138(2):882-97. doi: 10.1104/pp.105.062257. Epub 2005 May 27. Plant Physiol. 2005. PMID: 15923322 Free PMC article.
-
Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways.BMC Plant Biol. 2010 Jul 19;10:151. doi: 10.1186/1471-2229-10-151. BMC Plant Biol. 2010. PMID: 20642851 Free PMC article.
-
Antagonistic interaction between systemic acquired resistance and the abscisic acid-mediated abiotic stress response in Arabidopsis.Plant Cell. 2008 Jun;20(6):1678-92. doi: 10.1105/tpc.107.054296. Epub 2008 Jun 27. Plant Cell. 2008. PMID: 18586869 Free PMC article.
Cited by
-
Crosstalk between abscisic acid and nitric oxide under heat stress: exploring new vantage points.Plant Cell Rep. 2021 Aug;40(8):1429-1450. doi: 10.1007/s00299-021-02695-4. Epub 2021 Apr 28. Plant Cell Rep. 2021. PMID: 33909122 Review.
-
AtCYP710A1 gene-mediated stigmasterol production plays a role in imparting temperature stress tolerance in Arabidopsis thaliana.Plant Signal Behav. 2013 Feb;8(2):e23142. doi: 10.4161/psb.23142. Epub 2013 Jan 8. Plant Signal Behav. 2013. PMID: 23299431 Free PMC article.
-
Effects of Inorganic Salt Solutions on Vigour, Viability, Oxidative Metabolism and Germination Enzymes in Aged Cabbage and Lettuce Seeds.Plants (Basel). 2020 Sep 9;9(9):1164. doi: 10.3390/plants9091164. Plants (Basel). 2020. PMID: 32916793 Free PMC article.
-
Early Life Oxidative Stress and Long-Lasting Cardiovascular Effects on Offspring Conceived by Assisted Reproductive Technologies: A Review.Int J Mol Sci. 2020 Jul 22;21(15):5175. doi: 10.3390/ijms21155175. Int J Mol Sci. 2020. PMID: 32707756 Free PMC article. Review.
-
Oxidative stress tolerance in plants: novel interplay between auxin and reactive oxygen species signaling.Plant Signal Behav. 2013 Oct;8(10):doi: 10.4161/psb.25761. doi: 10.4161/psb.25761. Plant Signal Behav. 2013. PMID: 23887492 Free PMC article.
References
-
- Annamalai P, Yanaghiara S. Identification and characterization of a heat stress induced gene in cabbage encodes a kunitz type protease inhibitor. J Plant Physiol. 1999;155:226–233.
-
- Burke JJ. Identification of genetic diversity and mutations in higher plant acquired thermotolerance. Physiol Plant. 2001;112:167–170.
-
- Campbell AK, Trewavas AJ, Knight MR. Calcium imaging shows differential sensitivity to cooling and communication in luminous transgenic plants. Cell Calcium. 1996;19:211–218. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
