

Distinct role for microglia in rotenone-induced degeneration of dopaminergic neurons
- PMID: 11826108
- PMCID: PMC6758500
- DOI: 10.1523/JNEUROSCI.22-03-00782.2002
Distinct role for microglia in rotenone-induced degeneration of dopaminergic neurons
Abstract
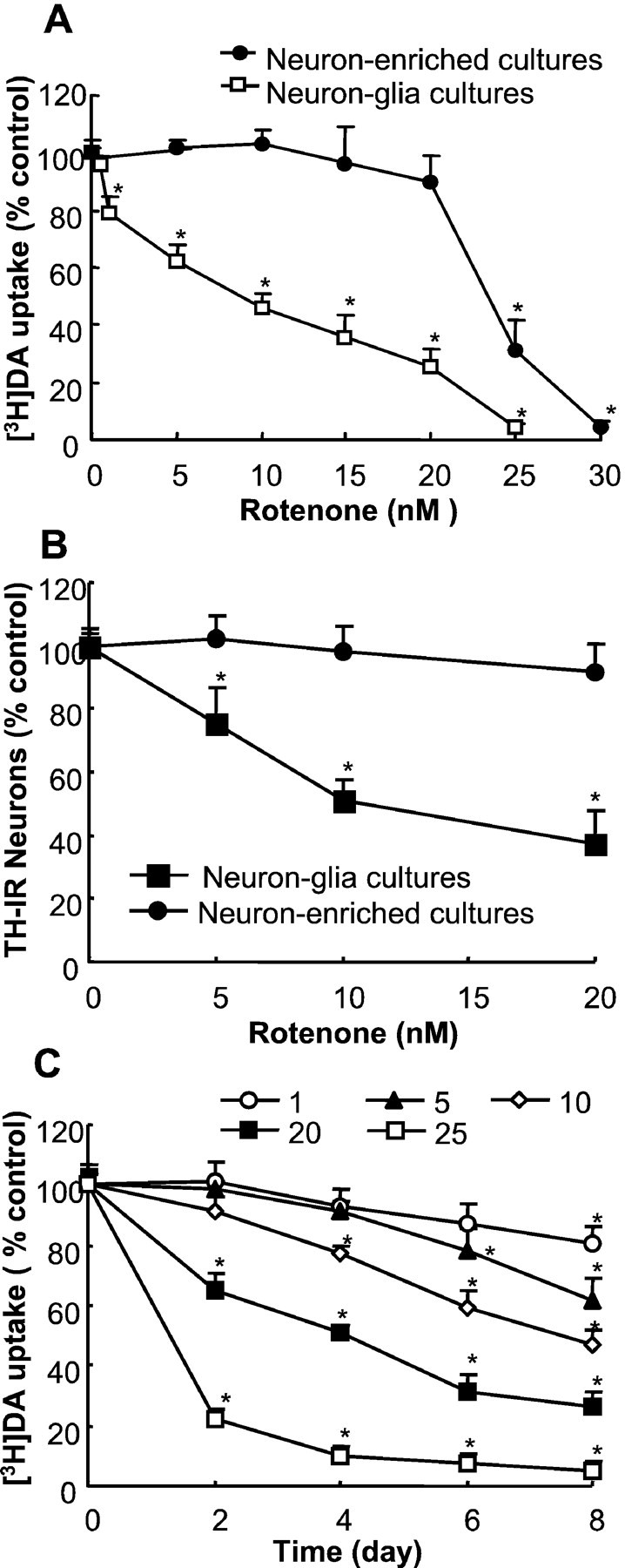
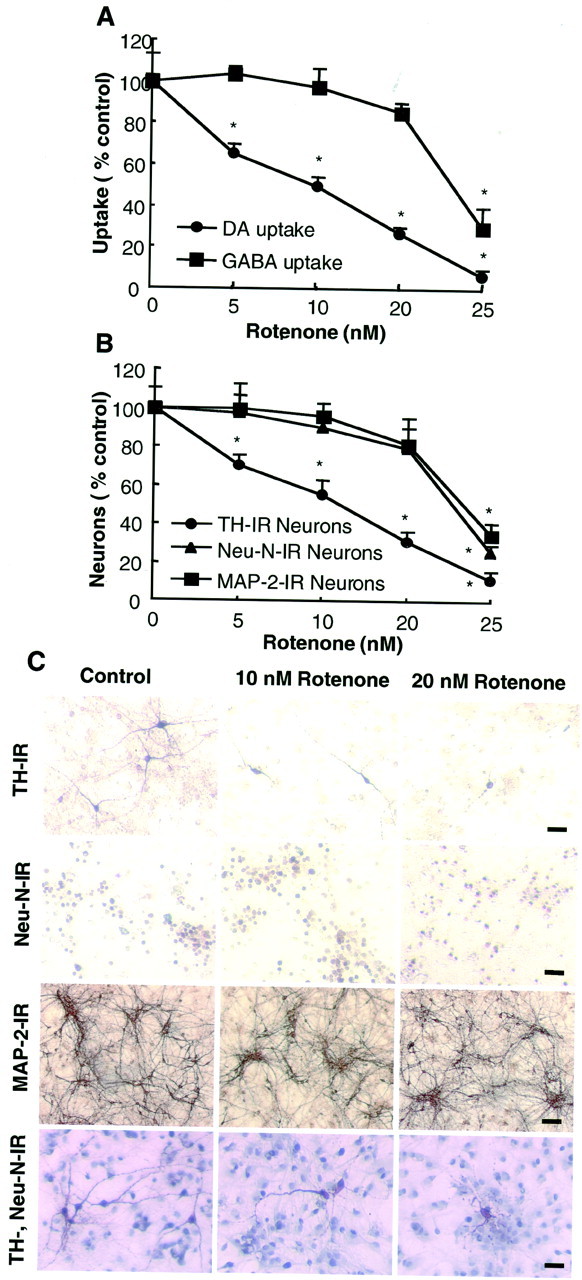
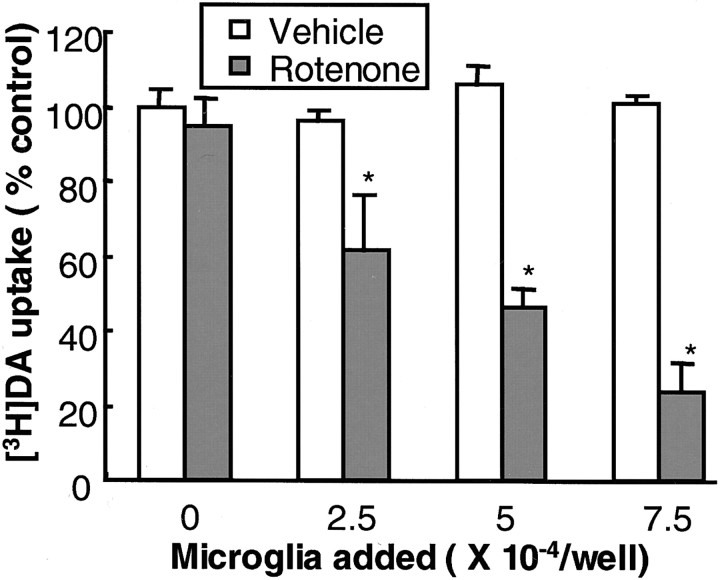
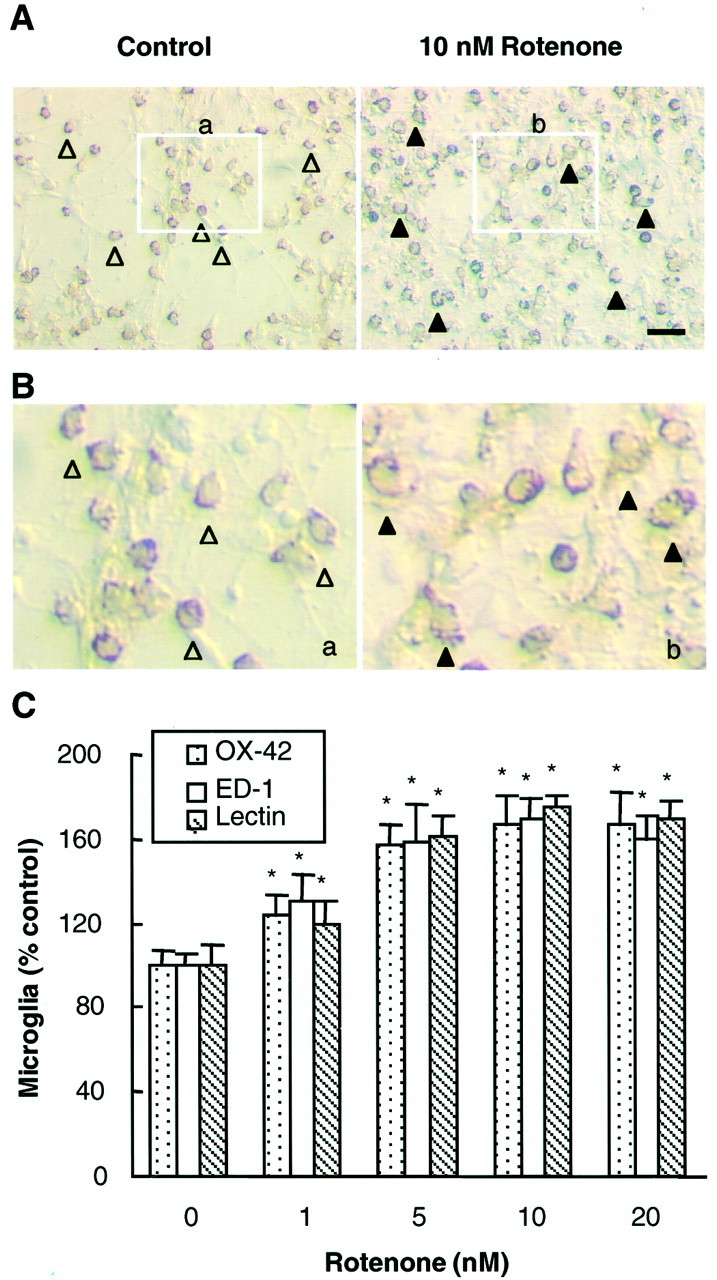
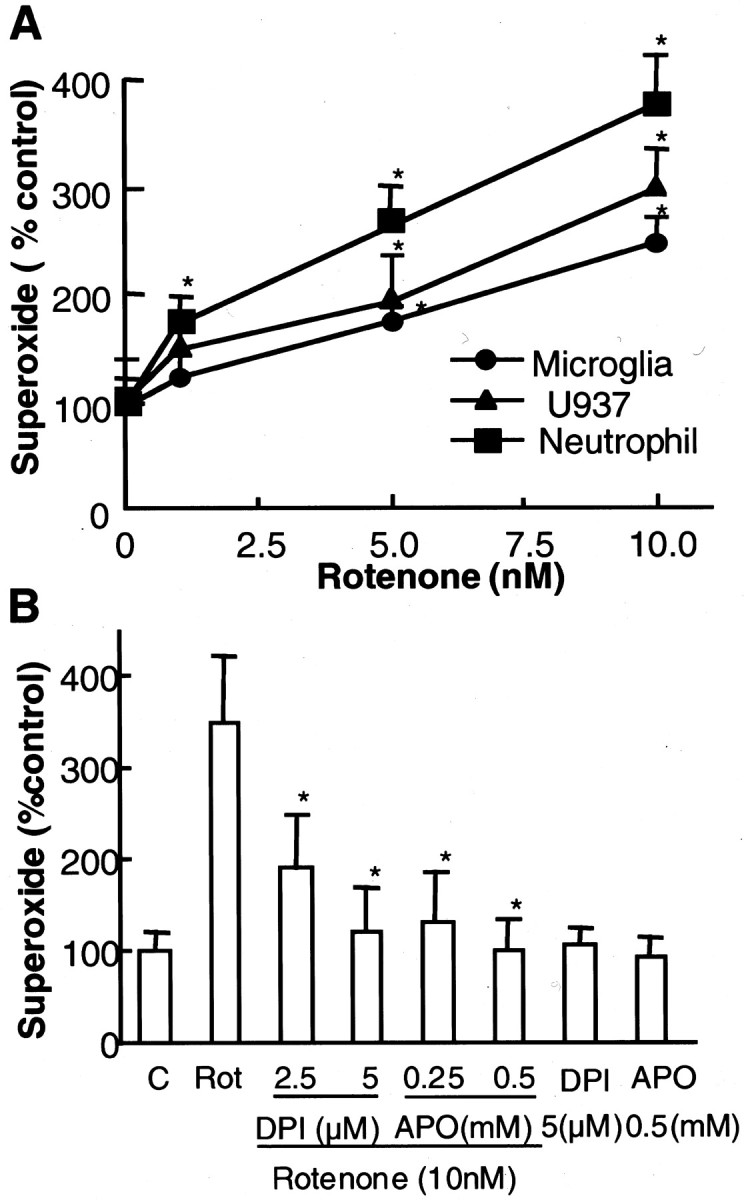
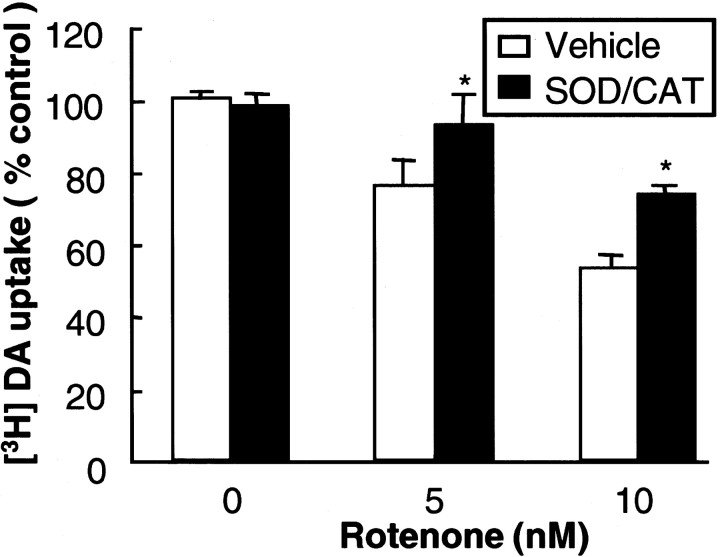
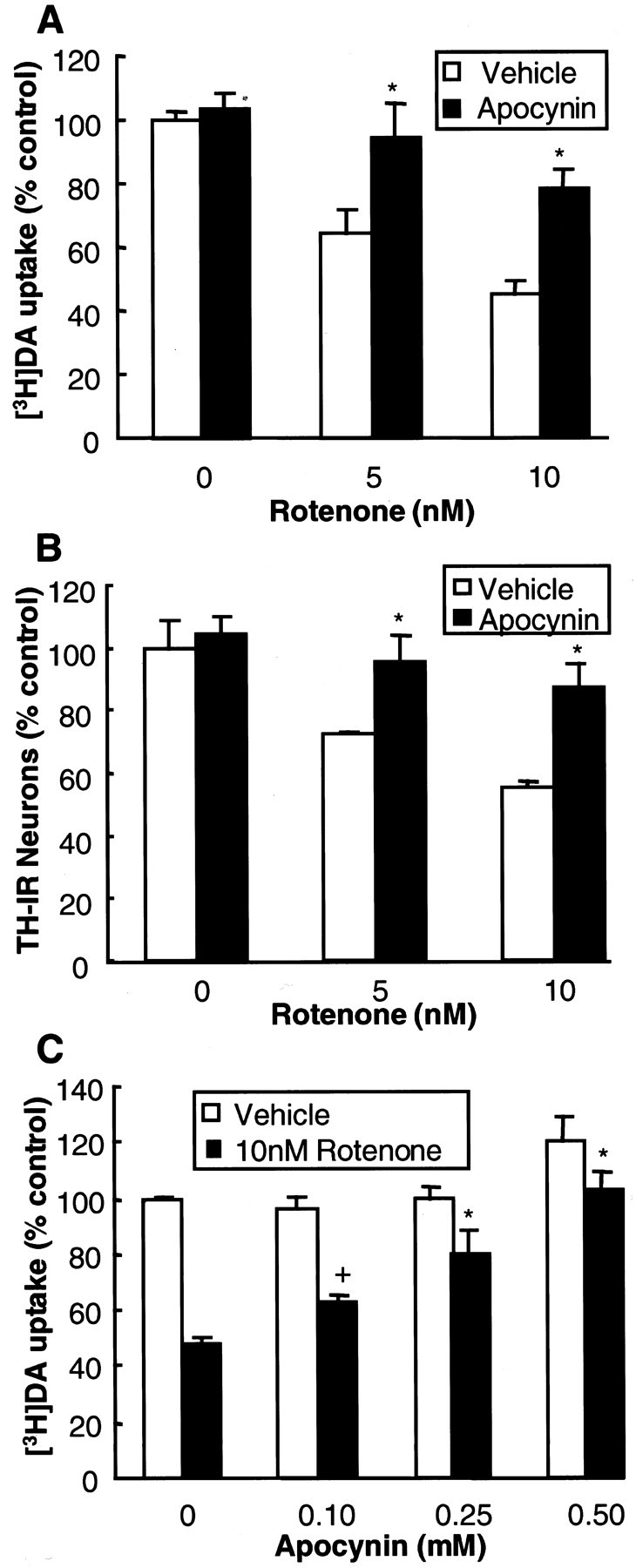
Increasing evidence has suggested an important role for environmental factors such as exposure to pesticides in the pathogenesis of Parkinson's disease. In experimental animals the exposure to a common herbicide, rotenone, induces features of parkinsonism; mechanistically, rotenone-induced destruction of dopaminergic neurons has been attributed to its inhibition of the activity of neuronal mitochondrial complex I. However, the role of microglia, the resident brain immune cells in rotenone-induced neurodegeneration, has not been reported. Using primary neuron-enriched and neuron/glia cultures from the rat mesencephalon, we discovered an extraordinary feature for rotenone-induced degeneration of cultured dopaminergic neurons. Although little neurotoxicity was detected in neuron-enriched cultures after treatment for 8 d with up to 20 nm rotenone, significant and selective dopaminergic neurodegeneration was observed in neuron/glia cultures 2 d after treatment with 20 nm rotenone or 8 d after treatment with 1 nm rotenone. The greatly enhanced neurodegenerative ability of rotenone was attributed to the presence of glia, especially microglia, because the addition of microglia to neuron-enriched cultures markedly increased their susceptibility to rotenone. Mechanistically, rotenone stimulated the release of superoxide from microglia that was attenuated by inhibitors of NADPH oxidase. Furthermore, inhibition of NADPH oxidase or scavenging of superoxide significantly reduced the rotenone-induced neurotoxicity. This is the first report demonstrating that microglia play a pivotal role in rotenone-induced degeneration of dopaminergic neurons. The results of this study should advance our understanding of the mechanism of action for pesticides in the pathogenesis of Parkinson's disease.
Figures
Similar articles
-
Critical role for microglial NADPH oxidase in rotenone-induced degeneration of dopaminergic neurons.J Neurosci. 2003 Jul 16;23(15):6181-7. doi: 10.1523/JNEUROSCI.23-15-06181.2003. J Neurosci. 2003. PMID: 12867501 Free PMC article.
-
Synergistic dopaminergic neurotoxicity of the pesticide rotenone and inflammogen lipopolysaccharide: relevance to the etiology of Parkinson's disease.J Neurosci. 2003 Feb 15;23(4):1228-36. doi: 10.1523/JNEUROSCI.23-04-01228.2003. J Neurosci. 2003. PMID: 12598611 Free PMC article.
-
Microglia enhance beta-amyloid peptide-induced toxicity in cortical and mesencephalic neurons by producing reactive oxygen species.J Neurochem. 2002 Nov;83(4):973-83. doi: 10.1046/j.1471-4159.2002.01210.x. J Neurochem. 2002. PMID: 12421370
-
Ultrafine carbon particles promote rotenone-induced dopamine neuronal loss through activating microglial NADPH oxidase.Toxicol Appl Pharmacol. 2017 May 1;322:51-59. doi: 10.1016/j.taap.2017.03.005. Epub 2017 Mar 7. Toxicol Appl Pharmacol. 2017. PMID: 28283350
-
HMGB1 acts on microglia Mac1 to mediate chronic neuroinflammation that drives progressive neurodegeneration.J Neurosci. 2011 Jan 19;31(3):1081-92. doi: 10.1523/JNEUROSCI.3732-10.2011. J Neurosci. 2011. PMID: 21248133 Free PMC article.
Cited by
-
Inflammatory cytokines in depression: neurobiological mechanisms and therapeutic implications.Neuroscience. 2013 Aug 29;246:199-229. doi: 10.1016/j.neuroscience.2013.04.060. Epub 2013 May 3. Neuroscience. 2013. PMID: 23644052 Free PMC article. Review.
-
Pesticides, microglial NOX2, and Parkinson's disease.J Biochem Mol Toxicol. 2013 Feb;27(2):137-49. doi: 10.1002/jbt.21464. Epub 2013 Jan 24. J Biochem Mol Toxicol. 2013. PMID: 23349115 Free PMC article.
-
Clozapine metabolites protect dopaminergic neurons through inhibition of microglial NADPH oxidase.J Neuroinflammation. 2016 May 16;13(1):110. doi: 10.1186/s12974-016-0573-z. J Neuroinflammation. 2016. PMID: 27184631 Free PMC article.
-
Role of secondary mediators in caffeine-mediated neuroprotection in maneb- and paraquat-induced Parkinson's disease phenotype in the mouse.Neurochem Res. 2012 Apr;37(4):875-84. doi: 10.1007/s11064-011-0682-0. Epub 2011 Dec 27. Neurochem Res. 2012. PMID: 22201039
-
Neuroimmunological processes in Parkinson's disease and their relation to α-synuclein: microglia as the referee between neuronal processes and peripheral immunity.ASN Neuro. 2013 Apr 30;5(2):113-39. doi: 10.1042/AN20120066. ASN Neuro. 2013. PMID: 23506036 Free PMC article. Review.
References
-
- Aloisi F. The role of microglia and astrocytes in CNS immune surveillance and immunopathology. Adv Exp Med Biol. 1999;468:123–133. - PubMed
-
- Babior BM. NADPH oxidase: an update. Blood. 1999;93:1464–1476. - PubMed
-
- Betarbet R, Sherer TB, MacKenzie G, Garcia-Osuna M, Panov AV, Greenamyre JT. Chronic systemic pesticide exposure reproduces features of Parkinson's disease. Nat Neurosci. 2000;3:1301–1306. - PubMed
-
- Cassarino DS, Fall CP, Swerdlow RH, Smith TS, Halvorsen EM, Miller SW, Parks JP, Parker WD, Jr, Bennett JP., Jr Elevated reactive oxygen species and antioxidant enzyme activities in animal and cellular models of Parkinson's disease. Biochim Biophys Acta. 1997;1362:77–86. - PubMed
-
- Chao CC, Hu S, Molitor TW, Shaskan EG, Peterson PK. Activated microglia mediate neuronal cell injury via a nitric oxide mechanism. J Immunol. 1992;149:2736–2741. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical