

Molecular determinants of species specificity in the coronavirus receptor aminopeptidase N (CD13): influence of N-linked glycosylation
- PMID: 11559807
- PMCID: PMC114546
- DOI: 10.1128/JVI.75.20.9741-9752.2001
Molecular determinants of species specificity in the coronavirus receptor aminopeptidase N (CD13): influence of N-linked glycosylation
Abstract
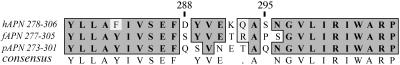
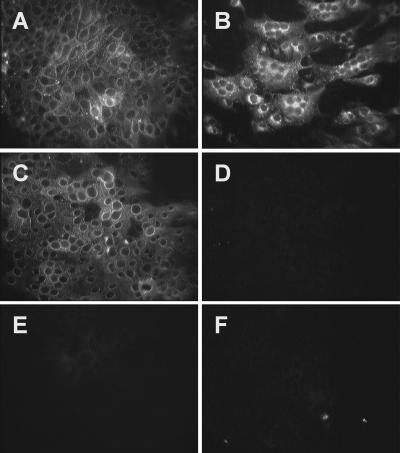
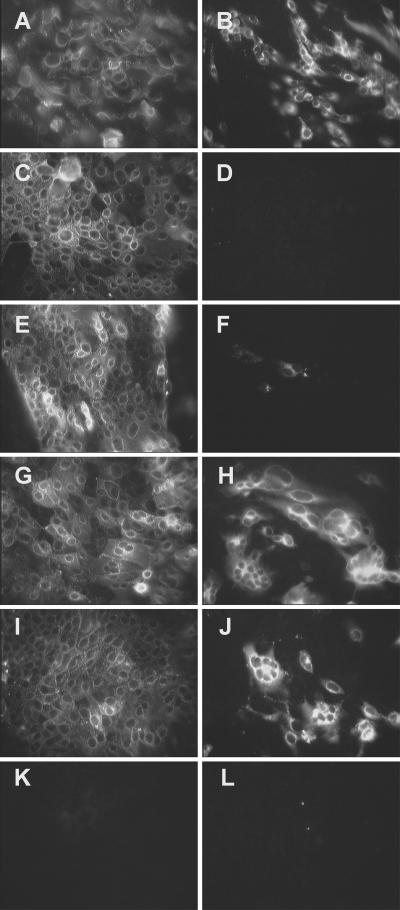
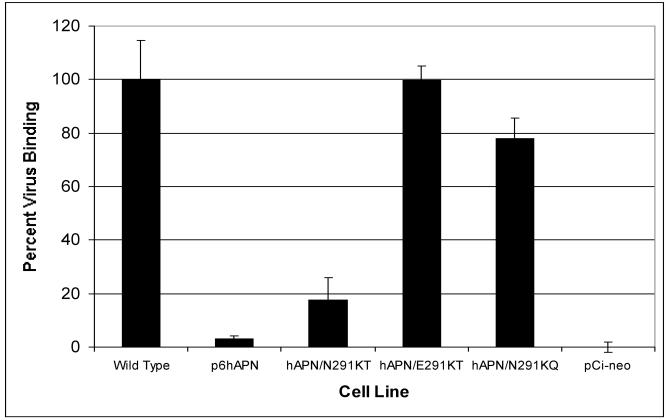
Aminopeptidase N (APN), a 150-kDa metalloprotease also called CD13, serves as a receptor for serologically related coronaviruses of humans (human coronavirus 229E [HCoV-229E]), pigs, and cats. These virus-receptor interactions can be highly species specific; for example, the human coronavirus can use human APN (hAPN) but not porcine APN (pAPN) as its cellular receptor, and porcine coronaviruses can use pAPN but not hAPN. Substitution of pAPN amino acids 283 to 290 into hAPN for the corresponding amino acids 288 to 295 introduced an N-glycosylation sequon at amino acids 291 to 293 that blocked HCoV-229E receptor activity of hAPN. Substitution of two amino acids that inserted an N-glycosylation site at amino acid 291 also resulted in a mutant hAPN that lacked receptor activity because it failed to bind HCoV-229E. Single amino acid revertants that removed this sequon at amino acids 291 to 293 but had one or five pAPN amino acid substitution(s) in this region all regained HCoV-229E binding and receptor activities. To determine if other N-linked glycosylation differences between hAPN, feline APN (fAPN), and pAPN account for receptor specificity of pig and cat coronaviruses, a mutant hAPN protein that, like fAPN and pAPN, lacked a glycosylation sequon at 818 to 820 was studied. This sequon is within the region that determines receptor activity for porcine and feline coronaviruses. Mutant hAPN lacking the sequon at amino acids 818 to 820 maintained HCoV-229E receptor activity but did not gain receptor activity for porcine or feline coronaviruses. Thus, certain differences in glycosylation between coronavirus receptors from different species are critical determinants in the species specificity of infection.
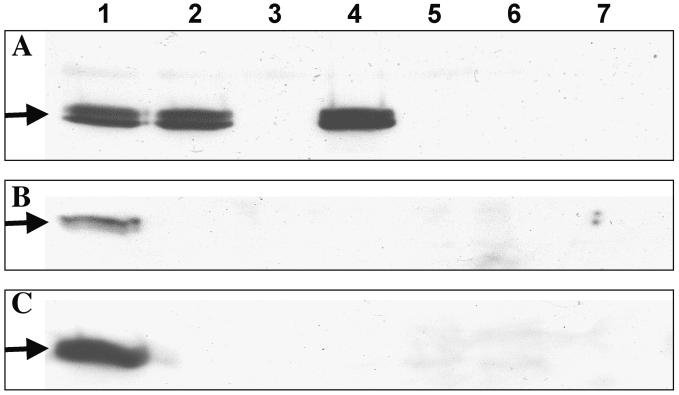
Figures
Similar articles
-
Mutational analysis of aminopeptidase N, a receptor for several group 1 coronaviruses, identifies key determinants of viral host range.J Virol. 2007 Feb;81(3):1261-73. doi: 10.1128/JVI.01510-06. Epub 2006 Nov 8. J Virol. 2007. PMID: 17093189 Free PMC article.
-
Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I.J Virol. 1996 Dec;70(12):8669-74. doi: 10.1128/JVI.70.12.8669-8674.1996. J Virol. 1996. PMID: 8970993 Free PMC article.
-
Characterization of functional domains in the human coronavirus HCV 229E receptor.J Gen Virol. 1996 Oct;77 ( Pt 10):2515-21. doi: 10.1099/0022-1317-77-10-2515. J Gen Virol. 1996. PMID: 8887485
-
Towards a coronavirus-based HIV multigene vaccine.Clin Dev Immunol. 2006 Jun-Dec;13(2-4):353-60. doi: 10.1080/17402520600579168. Clin Dev Immunol. 2006. PMID: 17162377 Free PMC article. Review.
-
Studying human pathogens in animal models: fine tuning the humanized mouse.Transgenic Res. 2005 Dec;14(6):803-6. doi: 10.1007/s11248-005-1676-y. Transgenic Res. 2005. PMID: 16315087 Free PMC article. Review.
Cited by
-
Characterization of a recombinant canine coronavirus with a distinct receptor-binding (S1) domain.Virology. 2012 Sep 1;430(2):90-9. doi: 10.1016/j.virol.2012.04.013. Epub 2012 May 18. Virology. 2012. PMID: 22609354 Free PMC article.
-
Genetic analysis of determinants for spike glycoprotein assembly into murine coronavirus virions: distinct roles for charge-rich and cysteine-rich regions of the endodomain.J Virol. 2004 Sep;78(18):9904-17. doi: 10.1128/JVI.78.18.9904-9917.2004. J Virol. 2004. PMID: 15331724 Free PMC article.
-
Targeting non-human coronaviruses to human cancer cells using a bispecific single-chain antibody.Gene Ther. 2005 Sep;12(18):1394-404. doi: 10.1038/sj.gt.3302535. Gene Ther. 2005. PMID: 15843808 Free PMC article.
-
Amino acid substitutions and an insertion in the spike glycoprotein extend the host range of the murine coronavirus MHV-A59.Virology. 2004 Jul 1;324(2):510-24. doi: 10.1016/j.virol.2004.04.005. Virology. 2004. PMID: 15207636 Free PMC article.
-
Genome organization and reverse genetic analysis of a type I feline coronavirus.J Virol. 2008 Feb;82(4):1851-9. doi: 10.1128/JVI.02339-07. Epub 2007 Dec 12. J Virol. 2008. PMID: 18077720 Free PMC article.
References
-
- Ashmun R A, Holmes K V, Shapiro L H, Favaloro E J, Razak K, de Crom R P G, Howard C J, Look A T. M3 CD13 (aminopeptidase N) cluster workshop report. 1. Leucocyte typing V. In: Schlossman S F, Boumsell L, Gilks W, Harlan J M, Kishimoto T, Morimoto C, Ritz J, Shaw S, Silverstein R, Springer T, Tedder T F, Todd R F, editors. White cell differentiation antigens. Proceedings of the Fifth International Workshop and Conference. Oxford, United Kingdom: Oxford University Press; 1995. pp. 771–775.
-
- Bandres J C, Wang Q F, O'Leary J, Baleaux F, Amara A, Hoxie J A, Zolla-Pazner S, Gorny M K. Human immunodeficiency virus (HIV) envelope binds to CXCR4 independently of CD4, and binding can be enhanced by interaction with soluble CD4 or by HIV envelope deglycosylation. J Virol. 1998;72:2500–2504. - PMC - PubMed
-
- Barnes K, Kenny A J, Turner A J. Localization of aminopeptidase N and dipeptidyl peptidase IV in pig striatum and in neuronal and glial cell cultures. Eur J Neurosci. 1994;6:531–537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
