

Distinct mesodermal signals, including BMPs from the septum transversum mesenchyme, are required in combination for hepatogenesis from the endoderm
- PMID: 11485993
- PMCID: PMC312750
- DOI: 10.1101/gad.904601
Distinct mesodermal signals, including BMPs from the septum transversum mesenchyme, are required in combination for hepatogenesis from the endoderm
Abstract
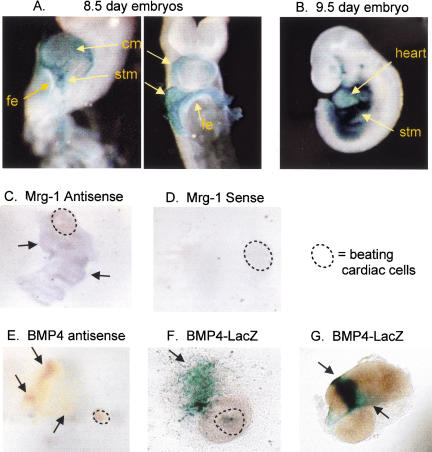
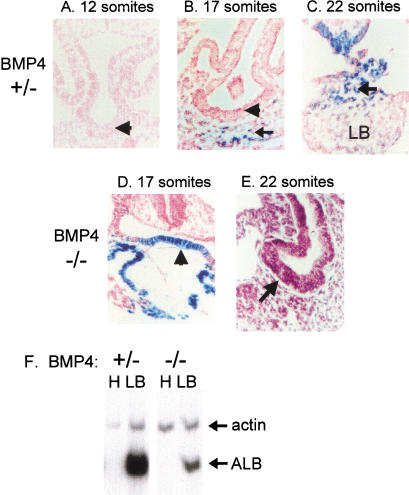
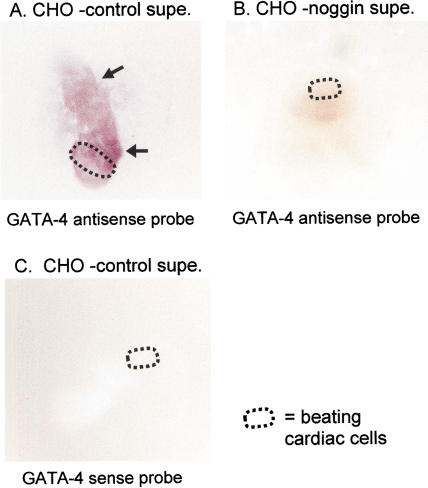
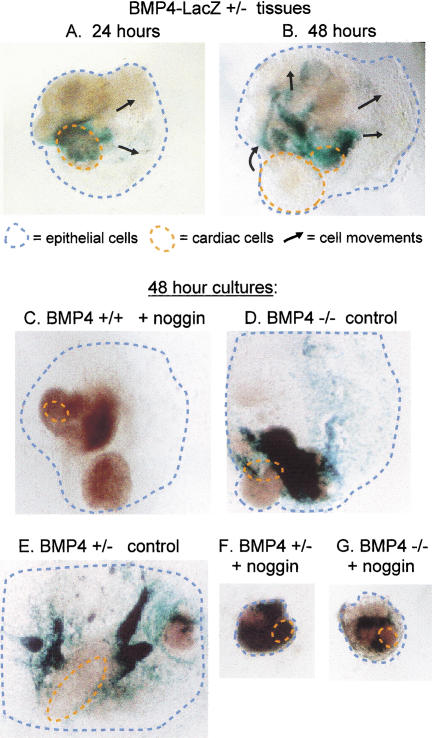
Mesodermal signaling is critical for patterning the embryonic endoderm into different tissue domains. Classical tissue transplant experiments in the chick and recent studies in the mouse indicated that interactions with the cardiogenic mesoderm are necessary and sufficient to induce the liver in the ventral foregut endoderm. Using molecular markers and functional assays, we now show that septum transversum mesenchyme cells, a distinct mesoderm cell type, are closely apposed to the ventral endoderm and contribute to hepatic induction. Specifically, using a mouse Bmp4 null mutation and an inhibitor of BMPs, we find that BMP signaling from the septum transversum mesenchyme is necessary to induce liver genes in the endoderm and to exclude a pancreatic fate. BMPs apparently function, in part, by affecting the levels of the GATA4 transcription factor, and work in parallel to FGF signaling from the cardiac mesoderm. BMP signaling also appears critical for morphogenetic growth of the hepatic endoderm into a liver bud. Thus, the endodermal domain for the liver is specified by simultaneous signaling from distinct mesodermal sources.
Figures



Similar articles
-
Gata4 expression in lateral mesoderm is downstream of BMP4 and is activated directly by Forkhead and GATA transcription factors through a distal enhancer element.Development. 2005 Aug;132(15):3405-17. doi: 10.1242/dev.01913. Epub 2005 Jun 29. Development. 2005. PMID: 15987774
-
Different thresholds of fibroblast growth factors pattern the ventral foregut into liver and lung.Development. 2005 Jan;132(1):35-47. doi: 10.1242/dev.01570. Epub 2004 Dec 2. Development. 2005. PMID: 15576401
-
A bipotential precursor population for pancreas and liver within the embryonic endoderm.Development. 2001 Mar;128(6):871-81. doi: 10.1242/dev.128.6.871. Development. 2001. PMID: 11222142
-
Teeth. Where and how to make them.Trends Genet. 1999 Feb;15(2):59-65. doi: 10.1016/s0168-9525(98)01662-x. Trends Genet. 1999. PMID: 10098408 Review.
-
Neural induction. A bird's eye view.Trends Genet. 1999 Jan;15(1):20-4. doi: 10.1016/s0168-9525(98)01620-5. Trends Genet. 1999. PMID: 10087929 Review.
Cited by
-
Transcriptome analysis reveals EBF1 ablation-induced injuries in cardiac system.Theranostics. 2024 Aug 12;14(12):4894-4915. doi: 10.7150/thno.92060. eCollection 2024. Theranostics. 2024. PMID: 39239522 Free PMC article.
-
Epigenetic modulation of cell fate during pancreas development.Trends Dev Biol. 2023;16:1-27. Trends Dev Biol. 2023. PMID: 38873037 Free PMC article.
-
Wnt/β-Catenin signaling pathway in hepatocellular carcinoma: pathogenic role and therapeutic target.Front Oncol. 2024 Apr 2;14:1367364. doi: 10.3389/fonc.2024.1367364. eCollection 2024. Front Oncol. 2024. PMID: 38634048 Free PMC article. Review.
-
Deletion of Tgm2 suppresses BMP-mediated hepatocyte-to-cholangiocyte metaplasia in ductular reaction.Cell Prolif. 2024 Oct;57(10):e13646. doi: 10.1111/cpr.13646. Epub 2024 Apr 16. Cell Prolif. 2024. PMID: 38623945 Free PMC article.
-
Portal Vein Thrombosis With Hypoplasia in the Left Lobe of the Liver: A Case Report.Cureus. 2024 Jan 25;16(1):e52964. doi: 10.7759/cureus.52964. eCollection 2024 Jan. Cureus. 2024. PMID: 38406154 Free PMC article.
References
-
- Andree B, Duprez D, Vorbusch B, Arnold HH, Brand T. BMP-2 induces ectopic expression of cardiac lineage markers and interferes with somite formation in chicken embryos. Mech Dev. 1998;70:119–131. - PubMed
-
- Barron M, Gao M, Lough J. Requirement for BMP and FGF signaling during cardiogenic induction in non-precardiac mesoderm is specific, transient, and cooperative. Dev Dyn. 2000;218:383–393. - PubMed
-
- Bei M, Maas R. FGFs and BMP4 induce both Msx1-independent and Msx1-dependent signaling pathways in early tooth development. Development. 1998;125:4325–4333. - PubMed
-
- Bossard P, Zaret KS. GATA transcription factors as potentiators of gut endoderm differentiation. Development. 1998;125:4909–4917. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases