

RNA is a structural element in retrovirus particles
- PMID: 11320254
- PMCID: PMC33195
- DOI: 10.1073/pnas.091000398
RNA is a structural element in retrovirus particles
Abstract
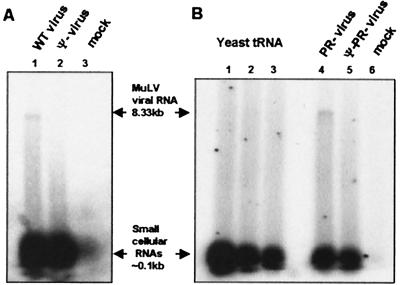
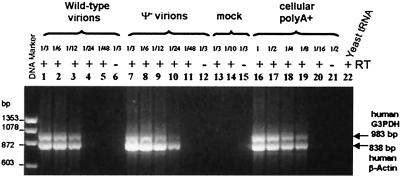
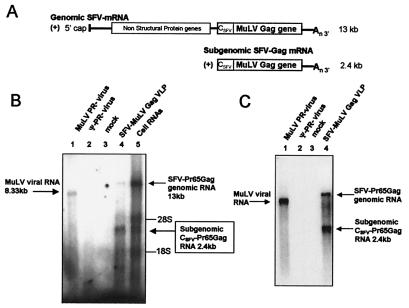
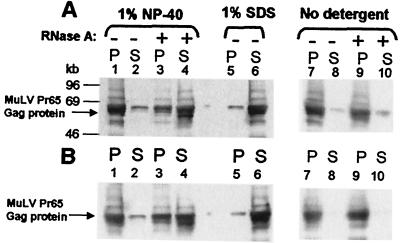
A single retroviral protein, Gag, is sufficient for virus particle assembly. While Gag is capable of specifically packaging the genomic RNA into the particle, this RNA species is unnecessary for particle assembly in vivo. In vitro, nucleic acids profoundly enhance the efficiency of assembly by recombinant Gag proteins, apparently by acting as "scaffolding" in the particle. To address the participation of RNA in retrovirus assembly in vivo, we analyzed murine leukemia virus particles that lack genomic RNA because of a deletion in the packaging signal of the viral RNA. We found that these particles contain cellular mRNA in place of genomic RNA. This result was particularly evident when Gag was expressed by using a Semliki Forest virus-derived vector: under these conditions, the Semliki Forest virus vector-directed mRNA became very abundant in the cells and was readily identified in the retroviral virus-like particles. Furthermore, we found that the retroviral cores were disrupted by treatment with RNase. Taken together, the data strongly suggest that RNA is a structural element in retrovirus particles.
Figures
Similar articles
-
Murine leukemia virus nucleocapsid mutant particles lacking viral RNA encapsidate ribosomes.J Virol. 2002 Nov;76(22):11405-13. doi: 10.1128/jvi.76.22.11405-11413.2002. J Virol. 2002. PMID: 12388701 Free PMC article.
-
The critical role of proximal gag sequences in feline immunodeficiency virus genome encapsidation.Virology. 2004 Sep 15;327(1):111-20. doi: 10.1016/j.virol.2004.06.014. Virology. 2004. PMID: 15327902
-
Semliki Forest virus-mediated production of retroviral vector RNA in retroviral packaging cells.Hum Gene Ther. 1997 Nov 20;8(17):2031-41. doi: 10.1089/hum.1997.8.17-2031. Hum Gene Ther. 1997. PMID: 9414252
-
Retroviral Gag protein-RNA interactions: Implications for specific genomic RNA packaging and virion assembly.Semin Cell Dev Biol. 2019 Feb;86:129-139. doi: 10.1016/j.semcdb.2018.03.015. Epub 2018 Apr 1. Semin Cell Dev Biol. 2019. PMID: 29580971 Free PMC article. Review.
-
Cell biology of retroviral RNA packaging.RNA Biol. 2011 Jul-Aug;8(4):572-80. doi: 10.4161/rna.8.4.16030. Epub 2011 Jul 1. RNA Biol. 2011. PMID: 21691151 Free PMC article. Review.
Cited by
-
Cross- and Co-Packaging of Retroviral RNAs and Their Consequences.Viruses. 2016 Oct 11;8(10):276. doi: 10.3390/v8100276. Viruses. 2016. PMID: 27727192 Free PMC article. Review.
-
A protein ballet around the viral genome orchestrated by HIV-1 reverse transcriptase leads to an architectural switch: from nucleocapsid-condensed RNA to Vpr-bridged DNA.Virus Res. 2013 Feb;171(2):287-303. doi: 10.1016/j.virusres.2012.09.008. Epub 2012 Sep 24. Virus Res. 2013. PMID: 23017337 Free PMC article.
-
Analysis of the contribution of cellular and viral RNA to the packaging of APOBEC3G into HIV-1 virions.Retrovirology. 2007 Jul 16;4:48. doi: 10.1186/1742-4690-4-48. Retrovirology. 2007. PMID: 17631688 Free PMC article.
-
Formation of the tRNALys packaging complex in HIV-1.FEBS Lett. 2010 Jan 21;584(2):359-65. doi: 10.1016/j.febslet.2009.11.038. FEBS Lett. 2010. PMID: 19914238 Free PMC article. Review.
-
An RNA structural switch regulates diploid genome packaging by Moloney murine leukemia virus.J Mol Biol. 2010 Feb 12;396(1):141-52. doi: 10.1016/j.jmb.2009.11.033. Epub 2009 Nov 17. J Mol Biol. 2010. PMID: 19931283 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
