

The Cia5 gene controls formation of the carbon concentrating mechanism in Chlamydomonas reinhardtii
- PMID: 11309511
- PMCID: PMC33211
- DOI: 10.1073/pnas.101534498
The Cia5 gene controls formation of the carbon concentrating mechanism in Chlamydomonas reinhardtii
Abstract
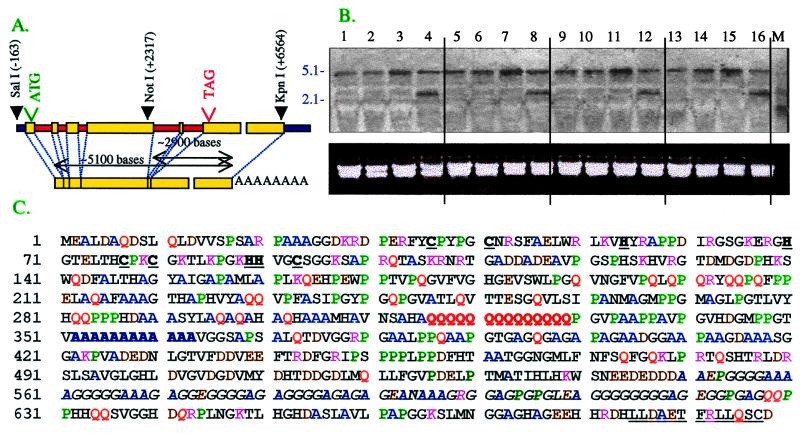
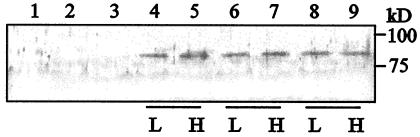
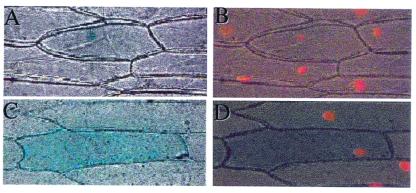
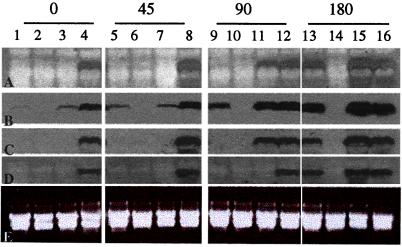
Wild-type Chlamydomonas reinhardtii cells shifted from high concentrations (5%) of CO2 to low, ambient levels (0.03%) rapidly increase transcription of mRNAs from several CO2-responsive genes. Simultaneously, they develop a functional carbon concentrating mechanism that allows the cells to greatly increase internal levels of CO2 and HCO3-. The cia5 mutant is defective in all of these phenotypes. A newly isolated gene, designated Cia5, restores transformed cia5 cells to the phenotype of wild-type cells. The 6,481-bp gene produces a 5.1-kb mRNA that is present constitutively in light in high and low CO2 both in wild-type cells and the cia5 mutant. It encodes a protein that has features of a putative transcription factor and that, likewise, is present constitutively in low and high CO2 conditions. Complementation of cia5 can be achieved with a truncated Cia5 gene that is missing the coding information for 54 C-terminal amino acids. Unlike wild-type cells or cia5 mutants transformed with an intact Cia5 gene, cia5 mutants complemented with the truncated gene exhibit constitutive synthesis of mRNAs from CO2-responsive genes in light under both high and low CO2 conditions. These discoveries suggest that posttranslational changes to the C-terminal domain control the ability of CIA5 to act as an inducer and directly or indirectly control transcription of CO2-responsive genes. Thus, CIA5 appears to be a master regulator of the carbon concentrating mechanism and is intimately involved in the signal transduction mechanism that senses and allows immediate responses to fluctuations in environmental CO2 and HCO3- concentrations.
Figures
Comment in
-
Acclimation of photosynthetic microorganisms to changing ambient CO2 concentration.Proc Natl Acad Sci U S A. 2001 Apr 24;98(9):4817-8. doi: 10.1073/pnas.101119898. Proc Natl Acad Sci U S A. 2001. PMID: 11320226 Free PMC article. No abstract available.
Similar articles
-
Ccm1, a regulatory gene controlling the induction of a carbon-concentrating mechanism in Chlamydomonas reinhardtii by sensing CO2 availability.Proc Natl Acad Sci U S A. 2001 Apr 24;98(9):5347-52. doi: 10.1073/pnas.081593498. Epub 2001 Apr 3. Proc Natl Acad Sci U S A. 2001. PMID: 11287669 Free PMC article.
-
The novel Myb transcription factor LCR1 regulates the CO2-responsive gene Cah1, encoding a periplasmic carbonic anhydrase in Chlamydomonas reinhardtii.Plant Cell. 2004 Jun;16(6):1466-77. doi: 10.1105/tpc.021162. Epub 2004 May 21. Plant Cell. 2004. PMID: 15155888 Free PMC article.
-
Glycolate production by a Chlamydomonas reinhardtii mutant lacking carbon-concentrating mechanism.J Biotechnol. 2021 Jul 20;335:39-46. doi: 10.1016/j.jbiotec.2021.06.009. Epub 2021 Jun 4. J Biotechnol. 2021. PMID: 34090947
-
Carbon-concentrating mechanism in a green alga, Chlamydomonas reinhardtii, revealed by transcriptome analyses.J Basic Microbiol. 2009 Feb;49(1):42-51. doi: 10.1002/jobm.200800352. J Basic Microbiol. 2009. PMID: 19253331 Review.
-
The Chlamydomonas CO2 -concentrating mechanism and its potential for engineering photosynthesis in plants.New Phytol. 2018 Jan;217(1):54-61. doi: 10.1111/nph.14749. Epub 2017 Aug 21. New Phytol. 2018. PMID: 28833179 Review.
Cited by
-
Co-expression networks in Chlamydomonas reveal significant rhythmicity in batch cultures and empower gene function discovery.Plant Cell. 2021 May 31;33(4):1058-1082. doi: 10.1093/plcell/koab042. Plant Cell. 2021. PMID: 33793846 Free PMC article.
-
The induction of pyrenoid synthesis by hyperoxia and its implications for the natural diversity of photosynthetic responses in Chlamydomonas.Elife. 2021 Dec 22;10:e67565. doi: 10.7554/eLife.67565. Elife. 2021. PMID: 34936552 Free PMC article.
-
Light and CO2/cAMP Signal Cross Talk on the Promoter Elements of Chloroplastic β-Carbonic Anhydrase Genes in the Marine Diatom Phaeodactylum tricornutum.Plant Physiol. 2016 Feb;170(2):1105-16. doi: 10.1104/pp.15.01738. Epub 2015 Dec 11. Plant Physiol. 2016. PMID: 26662605 Free PMC article.
-
Mitochondrial carbonic anhydrases are needed for optimal photosynthesis at low CO2 levels in Chlamydomonas.Plant Physiol. 2021 Nov 3;187(3):1387-1398. doi: 10.1093/plphys/kiab351. Plant Physiol. 2021. PMID: 34618049 Free PMC article.
-
Characterization of cooperative bicarbonate uptake into chloroplast stroma in the green alga Chlamydomonas reinhardtii.Proc Natl Acad Sci U S A. 2015 Jun 9;112(23):7315-20. doi: 10.1073/pnas.1501659112. Epub 2015 May 26. Proc Natl Acad Sci U S A. 2015. PMID: 26015566 Free PMC article.
References
-
- Raven J A. Can J Bot. 1991;69:908–924.
-
- Badger M R. In: The Biochemistry of Plants: A Comprehensive Treatise. Hatch M D, Boardman N K, editors. Vol. 10. New York: Academic; 1987. pp. 219–274.
-
- Kaplan A, Reinhold L. Annu Rev Plant Physiol Plant Mol Biol. 1999;50:539–570. - PubMed
-
- Spalding M H. In: The Molecular Biology of Chloroplasts and Mitochondria of Chlamydomonas. Rochaix J D, Goldschmidt-Clermont M, Merchant S, editors. Dordrecht, The Netherlands: Kluwer; 1998. pp. 529–547.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
