

Neuroprotection by encephalomyelitis: rescue of mechanically injured neurons and neurotrophin production by CNS-infiltrating T and natural killer cells
- PMID: 10884312
- PMCID: PMC6772346
- DOI: 10.1523/JNEUROSCI.20-14-05283.2000
Neuroprotection by encephalomyelitis: rescue of mechanically injured neurons and neurotrophin production by CNS-infiltrating T and natural killer cells
Abstract
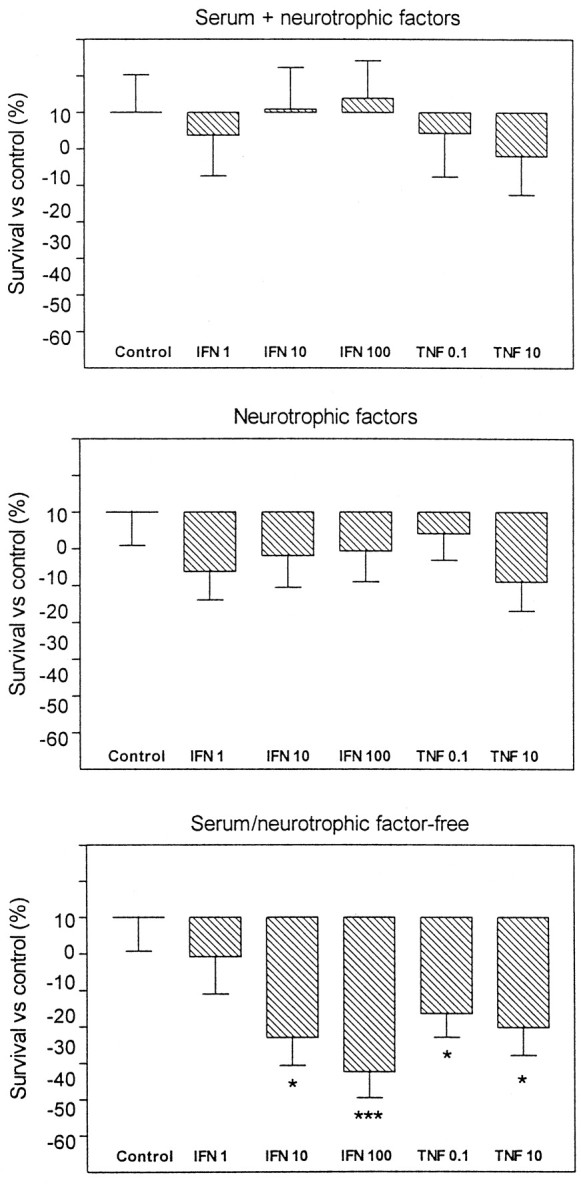
In experimental autoimmune encephalomyelitis (EAE), CD4(+) self-reactive T cells target myelin components of the CNS. However, the consequences of an autoaggressive T cell response against myelin for neurons are currently unknown. We herein demonstrate that EAE induced by active immunization with an encephalitogenic myelin basic protein peptide dramatically reduces the loss of spinal motoneurons after ventral root avulsion in rats. Both brain-derived neurotophic factor (BDNF)- and neurotrophin-3 (NT-3)-like immunoreactivities were detected in mainly T and natural killer (NK) cells in the spinal cord. In addition, very high levels of BDNF, NT-3, and glial cell line-derived neurotrophic factor mRNAs were present in T and NK cell populations infiltrating the CNS. Interestingly, bystander recruited NK and T cells displayed similar or higher neurotrophic factor levels compared with the EAE disease-driving encephalitogenic T cell population. High levels of tumor necrosis factor-alpha (TNF-alpha) and interferon-gamma (IFN-gamma) mRNAs were also detected, and both these cytokines can be harmful to several types of CNS cells, including neurons. However, treatment of embryonic motoneuron cultures with TNF-alpha or IFN-gamma only had a deleterious effect in cultures deprived of neurotrophic factors. These results suggest that the potentially neurodamaging consequences of severe CNS inflammation are curbed by the production of several potent neurotrophic factors in leukocytes.
Figures
Similar articles
-
Differential expression of neurotrophic factors and inflammatory cytokines by myelin basic protein-specific and other recruited T cells infiltrating the central nervous system during experimental autoimmune encephalomyelitis.Scand J Immunol. 2002 Mar;55(3):264-73. doi: 10.1046/j.0300-9475.2002.01038.x. Scand J Immunol. 2002. PMID: 11940233
-
Production of tumor necrosis factor-alpha as a result of glia-T-cell interaction correlates with the pathogenic activity of myelin basic protein-reactive T cells in experimental autoimmune encephalomyelitis.J Neurosci Res. 1996 Aug 15;45(4):400-9. doi: 10.1002/(SICI)1097-4547(19960815)45:4<400::AID-JNR9>3.0.CO;2-D. J Neurosci Res. 1996. PMID: 8872900
-
Innate IFN-γ promotes development of experimental autoimmune encephalomyelitis: a role for NK cells and M1 macrophages.Eur J Immunol. 2014 Oct;44(10):2903-17. doi: 10.1002/eji.201444612. Epub 2014 Aug 19. Eur J Immunol. 2014. PMID: 25056715
-
Function of neurotrophic factors beyond the nervous system: inflammation and autoimmune demyelination.Crit Rev Immunol. 2009;29(1):43-68. doi: 10.1615/critrevimmunol.v29.i1.20. Crit Rev Immunol. 2009. PMID: 19348610 Review.
-
The effects of interferon-gamma on the central nervous system.Mol Neurobiol. 1997 Feb-Apr;14(1-2):19-35. doi: 10.1007/BF02740619. Mol Neurobiol. 1997. PMID: 9170099 Free PMC article. Review.
Cited by
-
Genetic variability in the rat Aplec C-type lectin gene cluster regulates lymphocyte trafficking and motor neuron survival after traumatic nerve root injury.J Neuroinflammation. 2013 May 8;10:60. doi: 10.1186/1742-2094-10-60. J Neuroinflammation. 2013. PMID: 23656637 Free PMC article.
-
Impeded interaction between Schwann cells and axons in the absence of laminin alpha4.J Neurosci. 2005 Apr 6;25(14):3692-700. doi: 10.1523/JNEUROSCI.5225-04.2005. J Neurosci. 2005. PMID: 15814800 Free PMC article.
-
Neuroprotection and immunomodulation following intraspinal axotomy of motoneurons by treatment with adult mesenchymal stem cells.J Neuroinflammation. 2018 Aug 14;15(1):230. doi: 10.1186/s12974-018-1268-4. J Neuroinflammation. 2018. PMID: 30107848 Free PMC article.
-
Pathological CNS autoimmune disease triggered by traumatic spinal cord injury: implications for autoimmune vaccine therapy.J Neurosci. 2002 Apr 1;22(7):2690-700. doi: 10.1523/JNEUROSCI.22-07-02690.2002. J Neurosci. 2002. PMID: 11923434 Free PMC article.
-
Quantitative 1H magnetic resonance spectroscopic imaging determines therapeutic immunization efficacy in an animal model of Parkinson's disease.J Neurosci. 2005 Feb 16;25(7):1691-700. doi: 10.1523/JNEUROSCI.4364-04.2005. J Neurosci. 2005. PMID: 15716405 Free PMC article.
References
-
- Ben-Nun A, Wekerle H, Cohen IR. The rapid isolation of clonable antigen-specific T lymphocyte lines capable of mediating autoimmune encephalomyelitis. Eur J Immunol. 1981;11:195–199. - PubMed
-
- Besser M, Wank R. Cutting edge: clonally restricted production of the neurotrophins brain-derived neurotrophic factor and neurotrophin-3 mRNA by human immune cells and Th1/Th2-polarized expression of their receptors. J Immunol. 1999;162:6303–6306. - PubMed
-
- Braun A, Lommatzsch M, Mannsfeldt A, Neuhaus-Steinmetz U, Fischer A, Schnoy N, Lewin GR, Renz H. Cellular sources of enhanced brain-derived neurotrophic factor production in a mouse model of allergic inflammation. Am J Respir Cell Mol Biol. 1999;21:537–546. - PubMed
-
- Camu W, Henderson CE. Purification of embryonic rat motoneurons by panning on a monoclonal antibody to the low-affinity NGF receptor. J Neurosci Methods. 1992;44:59–70. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
