

Induction of apoptosis by Drosophila reaper, hid and grim through inhibition of IAP function
- PMID: 10675328
- PMCID: PMC305597
- DOI: 10.1093/emboj/19.4.589
Induction of apoptosis by Drosophila reaper, hid and grim through inhibition of IAP function
Abstract
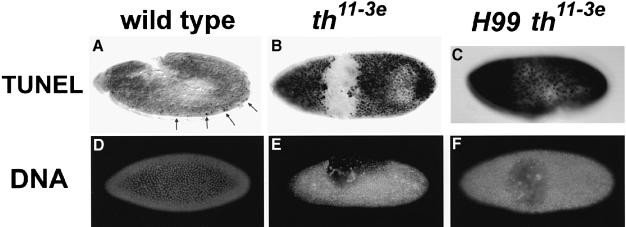
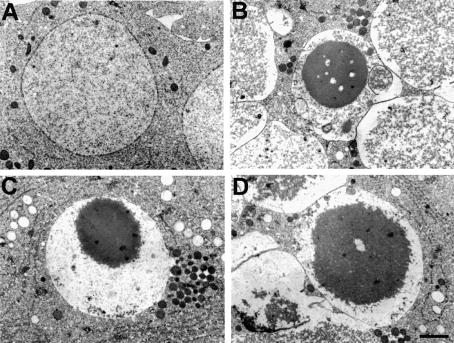
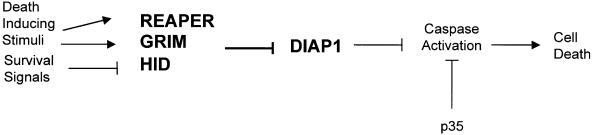
Induction of apoptosis in Drosophila requires the activity of three closely linked genes, reaper, hid and grim. Here we show that the proteins encoded by reaper, hid and grim activate cell death by inhibiting the anti-apoptotic activity of the Drosophila IAP1 (diap1) protein. In a genetic modifier screen, both loss-of-function and gain-of-function alleles in the endogenous diap1 gene were obtained, and the mutant proteins were functionally and biochemically characterized. Gain-of-function mutations in diap1 strongly suppressed reaper-, hid- and grim-induced apoptosis. Sequence analysis of these alleles revealed that they were caused by single amino acid changes in the baculovirus IAP repeat domains of diap1, a domain implicated in binding REAPER, HID and GRIM. Significantly, the corresponding mutant DIAP1 proteins displayed greatly reduced binding of REAPER, HID and GRIM, indicating that REAPER, HID and GRIM kill by forming a complex with DIAP1. These data provide strong in vivo evidence for a previously published model of cell death regulation in Drosophila.
Figures
Similar articles
-
Diverse domains of THREAD/DIAP1 are required to inhibit apoptosis induced by REAPER and HID in Drosophila.Genetics. 2000 Feb;154(2):669-78. doi: 10.1093/genetics/154.2.669. Genetics. 2000. PMID: 10655220 Free PMC article.
-
Distinct cell killing properties of the Drosophila reaper, head involution defective, and grim genes.Cell Death Differ. 1998 Nov;5(11):930-9. doi: 10.1038/sj.cdd.4400423. Cell Death Differ. 1998. PMID: 9846179
-
The Drosophila caspase inhibitor DIAP1 is essential for cell survival and is negatively regulated by HID.Cell. 1999 Aug 20;98(4):453-63. doi: 10.1016/s0092-8674(00)81974-1. Cell. 1999. PMID: 10481910
-
Cell death regulation by the mammalian IAP antagonist Diablo/Smac.Apoptosis. 2002 Apr;7(2):163-6. doi: 10.1023/a:1014318615955. Apoptosis. 2002. PMID: 11865200 Review.
-
Regulators of IAP function: coming to grips with the grim reaper.Curr Opin Cell Biol. 2003 Dec;15(6):717-24. doi: 10.1016/j.ceb.2003.10.002. Curr Opin Cell Biol. 2003. PMID: 14644196 Review.
Cited by
-
IAPs regulate the plasticity of cell migration by directly targeting Rac1 for degradation.EMBO J. 2012 Jan 4;31(1):14-28. doi: 10.1038/emboj.2011.423. Epub 2011 Nov 25. EMBO J. 2012. PMID: 22117219 Free PMC article.
-
A small-molecule ARTS mimetic promotes apoptosis through degradation of both XIAP and Bcl-2.Cell Death Dis. 2020 Jun 25;11(6):483. doi: 10.1038/s41419-020-2670-2. Cell Death Dis. 2020. PMID: 32587235 Free PMC article.
-
Temporal regulation of Drosophila IAP1 determines caspase functions in sensory organ development.J Cell Biol. 2009 Oct 19;187(2):219-31. doi: 10.1083/jcb.200905110. Epub 2009 Oct 12. J Cell Biol. 2009. PMID: 19822670 Free PMC article.
-
Reaper eliminates IAP proteins through stimulated IAP degradation and generalized translational inhibition.Nat Cell Biol. 2002 Jun;4(6):439-44. doi: 10.1038/ncb798. Nat Cell Biol. 2002. PMID: 12021770 Free PMC article.
-
Proteases for cell suicide: functions and regulation of caspases.Microbiol Mol Biol Rev. 2000 Dec;64(4):821-46. doi: 10.1128/MMBR.64.4.821-846.2000. Microbiol Mol Biol Rev. 2000. PMID: 11104820 Free PMC article. Review.
References
-
- Abrams J.M., White, K., Fessler, L.I. and Steller, H. (1993) Programmed cell death during Drosophila embryogenesis. Development, 117, 29–43. - PubMed
-
- Ambrosini G., Adida, C. and Altieri, D.C. (1997) A novel anti-apoptosis gene, Survivin, expressed in cancer and lymphoma. Nature Med., 3, 917–921. - PubMed
-
- Bergmann A., Agapite, J., McCall, K. and Steller, H. (1998a) The Drosophila gene hid is a direct molecular target of Ras-dependent survival signaling. Cell, 95, 331–341. - PubMed
-
- Bergmann A., Agapite, J. and Steller, H. (1998b) Mechanisms and control of programmed cell death in invertebrates. Oncogene, 17, 3215–3223. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
