

A novel snail-related transcription factor Smuc regulates basic helix-loop-helix transcription factor activities via specific E-box motifs
- PMID: 10606664
- PMCID: PMC102498
- DOI: 10.1093/nar/28.2.626
A novel snail-related transcription factor Smuc regulates basic helix-loop-helix transcription factor activities via specific E-box motifs
Abstract
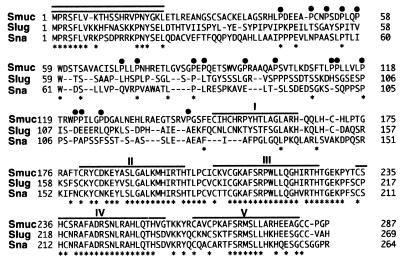
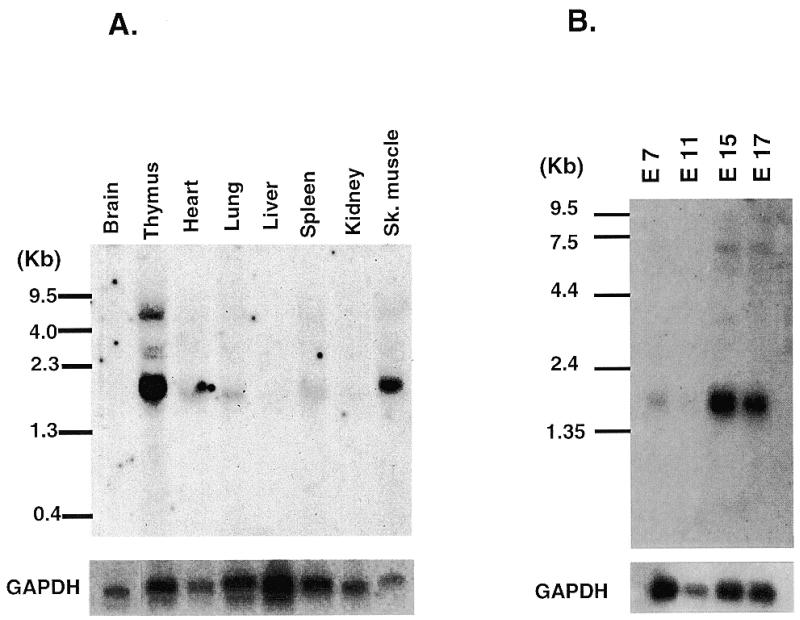
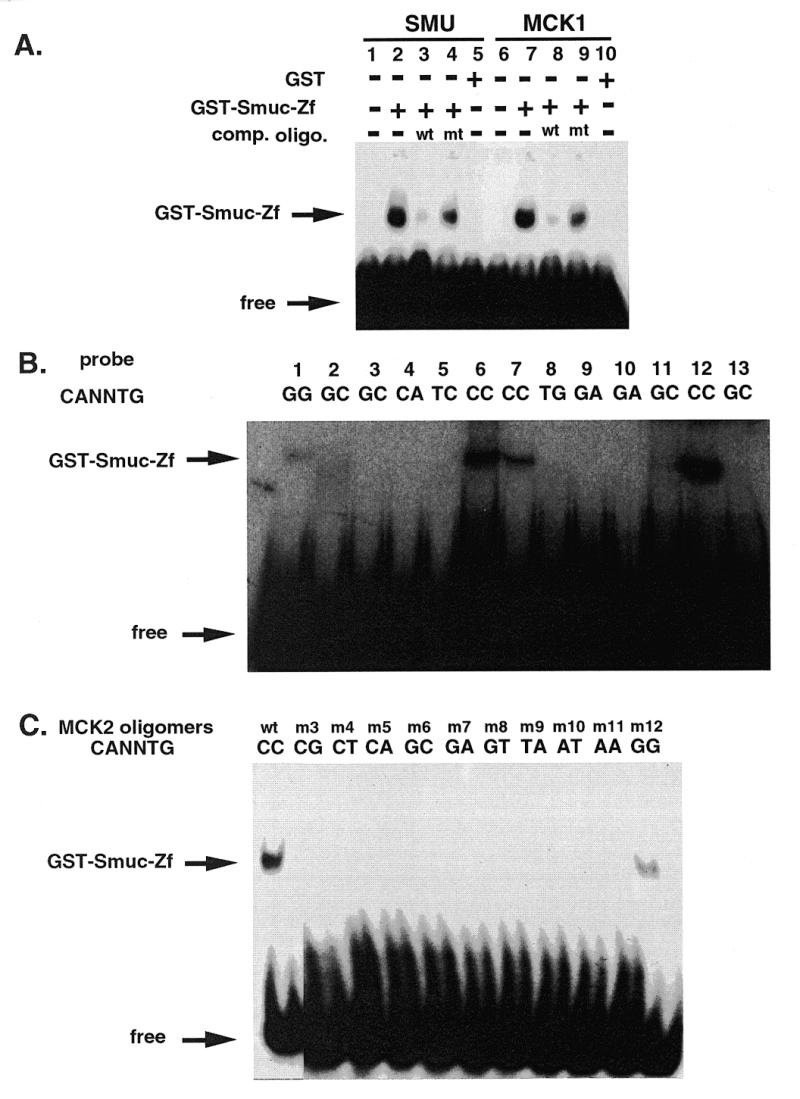
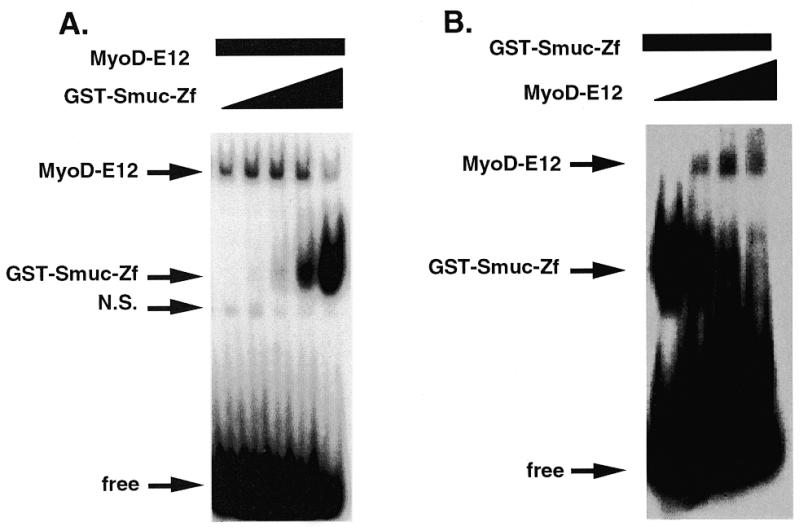
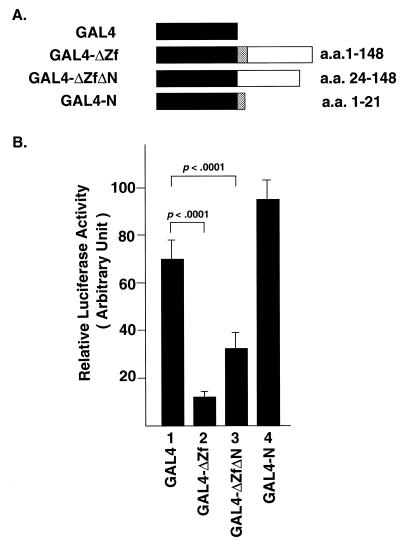
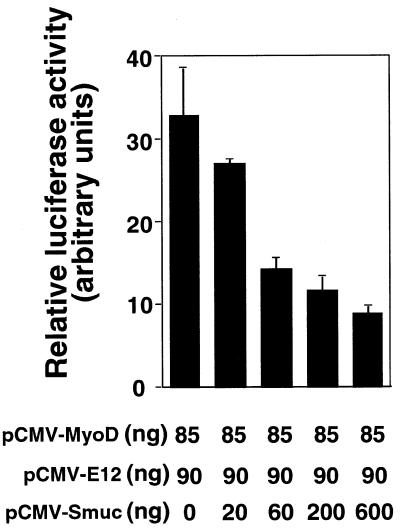
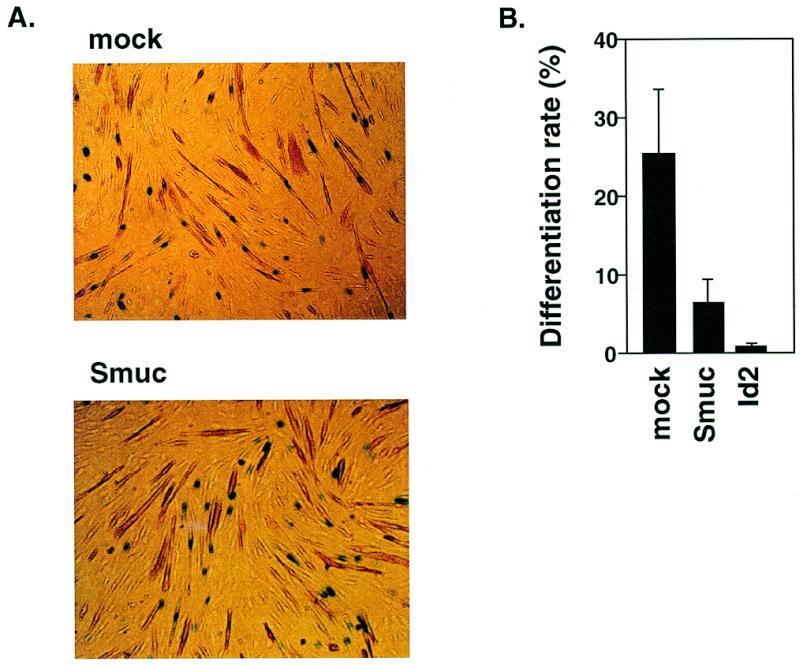
Snail family proteins are zinc finger transcriptional regulators first identified in Drosophila which play critical roles in cell fate determination. We identified a novel Snail -related gene from murine skeletalmusclecells designated Smuc. Northern blot analysis showed that Smuc was highly expressed in skeletal muscle and thymus. Smuc contains five putative DNA-binding zinc finger domains in its C-terminal half. In electrophoretic mobility shift assays, recombinant zinc finger domains of Smuc specifically bound to CAGGTG and CACCTG E-box motifs (CANNTG). Because basic helix-loop-helix transcription factors (bHLH) bind to the same E-box sequences, we examined whether Smuc competes with the myogenic bHLH factor MyoD for DNA binding. Smuc inhibited the binding of a MyoD-E12 complex to the CACCTG E-box sequence in a dose-dependent manner and suppressed the transcriptional activity of MyoD-E12. When heterologously targeted to the thymidine kinase promoter as fusion proteins with the GAL4 DNA-binding domain, the non-zinc finger domain of Smuc acted as a transcriptional repressor. Furthermore, overexpression of Smuc in myoblasts repressed transactivation of muscle differentiation marker Troponin T. Thus, Smuc might regulate bHLH transcription factors by zinc finger domains competing for E-box binding, and non-zinc finger repressor domains might also confer transcriptional repression to control differentiation processes.
Figures
Similar articles
-
A novel myogenic regulatory circuit controls slow/cardiac troponin C gene transcription in skeletal muscle.Mol Cell Biol. 1994 Mar;14(3):1870-85. doi: 10.1128/mcb.14.3.1870-1885.1994. Mol Cell Biol. 1994. PMID: 8114720 Free PMC article.
-
The basic helix-loop-helix transcription factor Mist1 functions as a transcriptional repressor of myoD.EMBO J. 1998 Mar 2;17(5):1412-22. doi: 10.1093/emboj/17.5.1412. EMBO J. 1998. PMID: 9482738 Free PMC article.
-
MyoR: a muscle-restricted basic helix-loop-helix transcription factor that antagonizes the actions of MyoD.Proc Natl Acad Sci U S A. 1999 Jan 19;96(2):552-7. doi: 10.1073/pnas.96.2.552. Proc Natl Acad Sci U S A. 1999. PMID: 9892671 Free PMC article.
-
Combinatorial control of muscle development by basic helix-loop-helix and MADS-box transcription factors.Proc Natl Acad Sci U S A. 1996 Sep 3;93(18):9366-73. doi: 10.1073/pnas.93.18.9366. Proc Natl Acad Sci U S A. 1996. PMID: 8790335 Free PMC article. Review.
-
Muscle differentiation: more complexity to the network of myogenic regulators.Curr Opin Genet Dev. 1998 Oct;8(5):539-44. doi: 10.1016/s0959-437x(98)80008-7. Curr Opin Genet Dev. 1998. PMID: 9794824 Review.
Cited by
-
Epithelial-mesenchymal plasticity (EMP) in wound healing: Exploring EMT mechanisms, regulatory network, and therapeutic opportunities.Heliyon. 2024 Jul 8;10(14):e34269. doi: 10.1016/j.heliyon.2024.e34269. eCollection 2024 Jul 30. Heliyon. 2024. PMID: 39108889 Free PMC article. Review.
-
Overexpression of Snai3 suppresses lymphoid- and enhances myeloid-cell differentiation.Eur J Immunol. 2012 Apr;42(4):1038-43. doi: 10.1002/eji.201142193. Eur J Immunol. 2012. PMID: 22531927 Free PMC article.
-
MicroRNA Regulation of Epithelial to Mesenchymal Transition.J Clin Med. 2016 Jan 14;5(1):8. doi: 10.3390/jcm5010008. J Clin Med. 2016. PMID: 26784241 Free PMC article. Review.
-
BPA Exposure Affects Mouse Gastruloids Axial Elongation by Perturbing the Wnt/β-Catenin Pathway.Int J Mol Sci. 2024 Jul 19;25(14):7924. doi: 10.3390/ijms25147924. Int J Mol Sci. 2024. PMID: 39063166 Free PMC article.
-
Snail3 orthologues in vertebrates: divergent members of the Snail zinc-finger gene family.Dev Genes Evol. 2004 Jan;214(1):47-53. doi: 10.1007/s00427-003-0373-1. Epub 2003 Dec 4. Dev Genes Evol. 2004. PMID: 14655014
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
