

Targeted recombination demonstrates that the spike gene of transmissible gastroenteritis coronavirus is a determinant of its enteric tropism and virulence
- PMID: 10438851
- PMCID: PMC104288
- DOI: 10.1128/JVI.73.9.7607-7618.1999
Targeted recombination demonstrates that the spike gene of transmissible gastroenteritis coronavirus is a determinant of its enteric tropism and virulence
Abstract
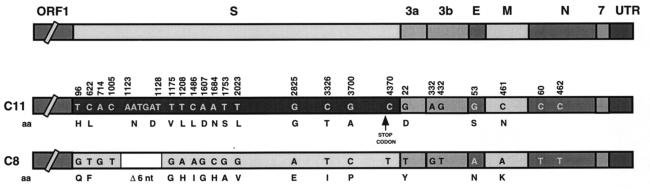
Targeted recombination within the S (spike) gene of transmissible gastroenteritis coronavirus (TGEV) was promoted by passage of helper respiratory virus isolates in cells transfected with a TGEV-derived defective minigenome carrying the S gene from an enteric isolate. The minigenome was efficiently replicated in trans and packaged by the helper virus, leading to the formation of true recombinant and pseudorecombinant viruses containing the S proteins of both enteric and respiratory TGEV strains in their envelopes. The recombinants acquired an enteric tropism, and their analysis showed that they were generated by homologous recombination that implied a double crossover in the S gene resulting in replacement of most of the respiratory, attenuated strain S gene (nucleotides 96 to 3700) by the S gene of the enteric, virulent isolate. The recombinant virus was virulent and rapidly evolved in swine testis cells by the introduction of point mutations and in-phase codon deletions in a domain of the S gene (nucleotides 217 to 665) previously implicated in the tropism of TGEV. The helper virus, with an original respiratory tropism, was also found in the enteric tract, probably because pseudorecombinant viruses carrying the spike proteins from the respiratory strain and the enteric virus in their envelopes were formed. These results demonstrated that a change in the tropism and virulence of TGEV can be engineered by sequence changes in the S gene.
Figures
 ) or the ileum (░⃞) of piglets infected with the strain
PTV plus the minigenome M54.
) or the ileum (░⃞) of piglets infected with the strain
PTV plus the minigenome M54.
Similar articles
-
Two amino acid changes at the N-terminus of transmissible gastroenteritis coronavirus spike protein result in the loss of enteric tropism.Virology. 1997 Jan 20;227(2):378-88. doi: 10.1006/viro.1996.8344. Virology. 1997. PMID: 9018137 Free PMC article.
-
Molecular bases of tropism in the PUR46 cluster of transmissible gastroenteritis coronaviruses.Adv Exp Med Biol. 1995;380:557-62. doi: 10.1007/978-1-4615-1899-0_89. Adv Exp Med Biol. 1995. PMID: 8830541
-
The spike protein of transmissible gastroenteritis coronavirus controls the tropism of pseudorecombinant virions engineered using synthetic minigenomes.Adv Exp Med Biol. 1998;440:207-14. doi: 10.1007/978-1-4615-5331-1_25. Adv Exp Med Biol. 1998. PMID: 9782282
-
An overview of immunological and genetic methods for detecting swine coronaviruses, transmissible gastroenteritis virus, and porcine respiratory coronavirus in tissues.Adv Exp Med Biol. 1997;412:37-46. doi: 10.1007/978-1-4899-1828-4_4. Adv Exp Med Biol. 1997. PMID: 9191988 Review.
-
Coronavirus reverse genetics and development of vectors for gene expression.Curr Top Microbiol Immunol. 2005;287:161-97. doi: 10.1007/3-540-26765-4_6. Curr Top Microbiol Immunol. 2005. PMID: 15609512 Free PMC article. Review.
Cited by
-
Long-distance RNA-RNA interactions in the coronavirus genome form high-order structures promoting discontinuous RNA synthesis during transcription.J Virol. 2013 Jan;87(1):177-86. doi: 10.1128/JVI.01782-12. Epub 2012 Oct 10. J Virol. 2013. PMID: 23055566 Free PMC article.
-
Tissue and cellular tropism of the coronavirus associated with severe acute respiratory syndrome: an in-situ hybridization study of fatal cases.J Pathol. 2004 Feb;202(2):157-63. doi: 10.1002/path.1510. J Pathol. 2004. PMID: 14743497 Free PMC article.
-
Development and efficacy of a novel live-attenuated QX-like nephropathogenic infectious bronchitis virus vaccine in China.Vaccine. 2015 Feb 25;33(9):1113-20. doi: 10.1016/j.vaccine.2015.01.036. Epub 2015 Jan 27. Vaccine. 2015. PMID: 25636916 Free PMC article.
-
Molecular Evolution of Human Coronavirus Genomes.Trends Microbiol. 2017 Jan;25(1):35-48. doi: 10.1016/j.tim.2016.09.001. Epub 2016 Oct 19. Trends Microbiol. 2017. PMID: 27743750 Free PMC article. Review.
-
Bioinformatic prediction of immunodominant regions in spike protein for early diagnosis of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2).PeerJ. 2021 Apr 8;9:e11232. doi: 10.7717/peerj.11232. eCollection 2021. PeerJ. 2021. PMID: 33889450 Free PMC article.
References
-
- Bergelson J M, Cunningham J A, Droguett G, Kurt-Jones E A, Krithivas A, Hong J S, Horwitz M S, Crowell R L, Finberg R W. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science. 1997;275:1320–1323. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
