

Tyrosine phosphorylation of A17 during vaccinia virus infection: involvement of the H1 phosphatase and the F10 kinase
- PMID: 10438817
- PMCID: PMC104254
- DOI: 10.1128/JVI.73.9.7287-7296.1999
Tyrosine phosphorylation of A17 during vaccinia virus infection: involvement of the H1 phosphatase and the F10 kinase
Abstract
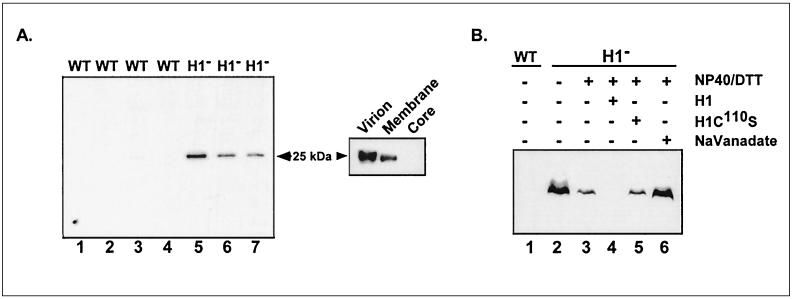
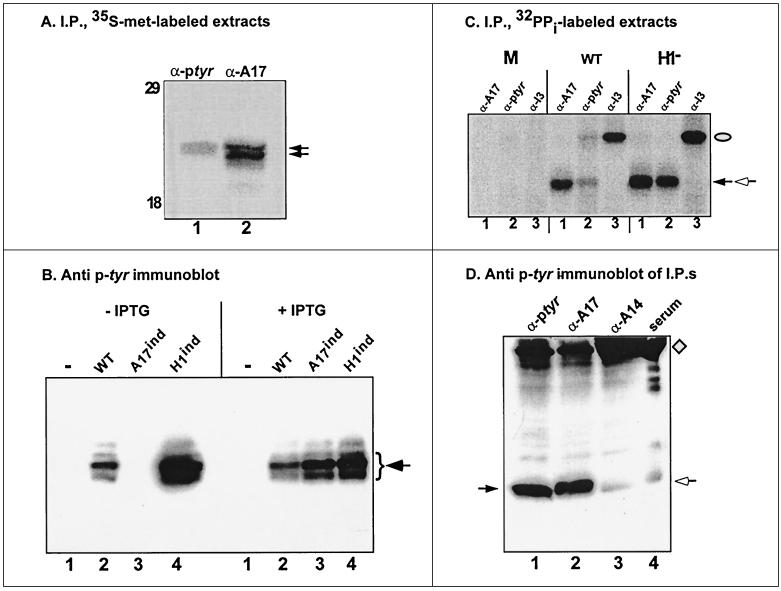
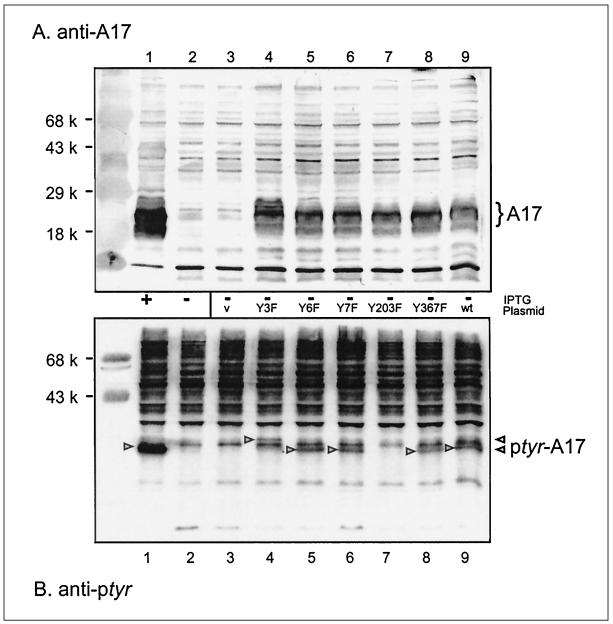
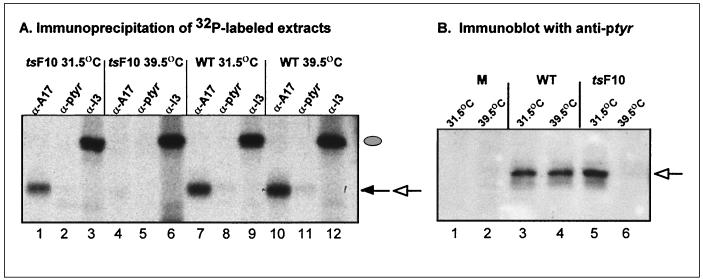
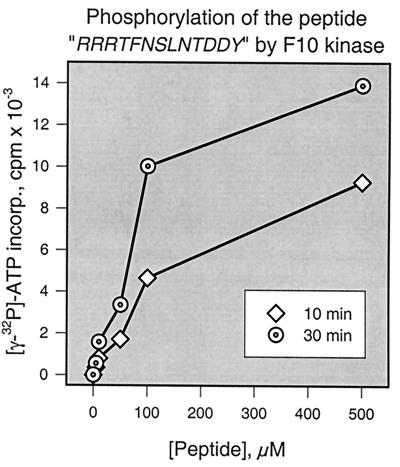
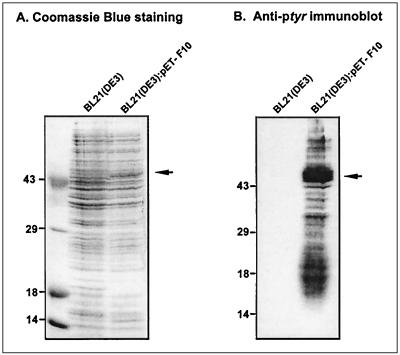
Vaccinia virus encodes two protein kinases (B1 and F10) and a dual-specificity phosphatase (VH1), suggesting that phosphorylation and dephosphorylation of substrates on serine/threonine and tyrosine residues are important in regulating diverse aspects of the viral life cycle. Using a recombinant in which expression of the H1 phosphatase can be regulated experimentally (vindH1), we have previously demonstrated that repression of H1 leads to the maturation of noninfectious virions that contain several hyperphosphorylated substrates (K. Liu et al., J. Virol. 69:7823-7834). In this report, we demonstrate that among these is a 25-kDa protein that is phosphorylated on tyrosine residues in H1-deficient virions and can be dephosphorylated by recombinant H1. We demonstrate that the 25-kDa phosphoprotein represents the product of the A17 gene and that A17 is phosphorylated on serine, threonine, and tyrosine residues during infection. Detection of phosphotyrosine within A17 is abrogated when Tyr(203) (but not Tyr(3), Tyr(6), or Tyr(7)) is mutated to phenylalanine, suggesting strongly that this amino acid is the site of tyrosine phosphorylation. Phosphorylation of A17 fails to occur during nonpermissive infections performed with temperature-sensitive mutants defective in the F10 kinase. Our data suggest that this enzyme, which was initially characterized as a serine/threonine kinase, might in fact have dual specificity. This hypothesis is strengthened by the observation that Escherichia coli induced to express F10 contain multiple proteins which are recognized by antiphosphotyrosine antiserum. This study presents the first evidence for phosphotyrosine signaling during vaccinia virus infection and implicates the F10 kinase and the H1 phosphatase as the dual-specificity enzymes that direct this cycle of reversible phosphorylation.
Figures
Similar articles
-
Elucidating the essential role of the A14 phosphoprotein in vaccinia virus morphogenesis: construction and characterization of a tetracycline-inducible recombinant.J Virol. 2000 Apr;74(8):3682-95. doi: 10.1128/jvi.74.8.3682-3695.2000. J Virol. 2000. PMID: 10729144 Free PMC article.
-
Physical and functional interactions between vaccinia virus F10 protein kinase and virion assembly proteins A30 and G7.J Virol. 2004 Jan;78(1):266-74. doi: 10.1128/jvi.78.1.266-274.2004. J Virol. 2004. PMID: 14671108 Free PMC article.
-
Vaccinia virus gene H5R encodes a protein that is phosphorylated by the multisubstrate vaccinia virus B1R protein kinase.J Virol. 1995 Mar;69(3):1819-26. doi: 10.1128/JVI.69.3.1819-1826.1995. J Virol. 1995. PMID: 7853522 Free PMC article.
-
Serine/threonine/tyrosine phosphorylation regulates DNA binding of bacterial transcriptional regulators.Microbiology (Reading). 2015 Sep;161(9):1720-1729. doi: 10.1099/mic.0.000148. Epub 2015 Jul 23. Microbiology (Reading). 2015. PMID: 26220449 Review.
-
Importance of Tyrosine Phosphorylation in Hormone-Regulated Plant Growth and Development.Int J Mol Sci. 2022 Jun 13;23(12):6603. doi: 10.3390/ijms23126603. Int J Mol Sci. 2022. PMID: 35743047 Free PMC article. Review.
Cited by
-
The vaccinia virus A33R protein provides a chaperone function for viral membrane localization and tyrosine phosphorylation of the A36R protein.J Virol. 2001 Jan;75(1):303-10. doi: 10.1128/JVI.75.1.303-310.2001. J Virol. 2001. PMID: 11119600 Free PMC article.
-
Elucidating the essential role of the A14 phosphoprotein in vaccinia virus morphogenesis: construction and characterization of a tetracycline-inducible recombinant.J Virol. 2000 Apr;74(8):3682-95. doi: 10.1128/jvi.74.8.3682-3695.2000. J Virol. 2000. PMID: 10729144 Free PMC article.
-
Cell biological and functional characterization of the vaccinia virus F10 kinase: implications for the mechanism of virion morphogenesis.J Virol. 2005 Feb;79(4):2171-90. doi: 10.1128/JVI.79.4.2171-2190.2005. J Virol. 2005. PMID: 15681420 Free PMC article.
-
Participation of vaccinia virus l2 protein in the formation of crescent membranes and immature virions.J Virol. 2011 Mar;85(6):2504-11. doi: 10.1128/JVI.02505-10. Epub 2011 Jan 12. J Virol. 2011. PMID: 21228235 Free PMC article.
-
Phosphorylation meets nuclear import: a review.Cell Commun Signal. 2010 Dec 23;8:32. doi: 10.1186/1478-811X-8-32. Cell Commun Signal. 2010. PMID: 21182795 Free PMC article.
References
-
- Banham A H, Smith G L. Vaccinia virus gene B1R encodes a 34-kDa serine/threonine protein kinase that localizes in cytoplasmic factories and is packaged into virions. Virology. 1992;191:803–812. - PubMed
-
- Beaud G, Sharif A, Topa-Masse A, Leader D P. Ribosomal protein S2/Sa kinase purified from HeLa cells infected with vaccinia virus corresponds to the B1R protein kinase and phosphorylates in vitro the viral ssDNA-binding protein. J Gen Virol. 1994;75:283–293. - PubMed
-
- Casnellie J E. Assay of protein kinases using peptides with basic residues for phosphocellulose binding. Methods Enzymol. 1991;200:115–121. - PubMed
-
- Condit R C, Motyczka A, Spizz G. Isolation, characterization and physical mapping of temperature sensitive mutants of vaccinia virus. Virology. 1983;128:429–443. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
