

A GATA-2/estrogen receptor chimera functions as a ligand-dependent negative regulator of self-renewal
- PMID: 10421636
- PMCID: PMC316885
- DOI: 10.1101/gad.13.14.1847
A GATA-2/estrogen receptor chimera functions as a ligand-dependent negative regulator of self-renewal
Abstract
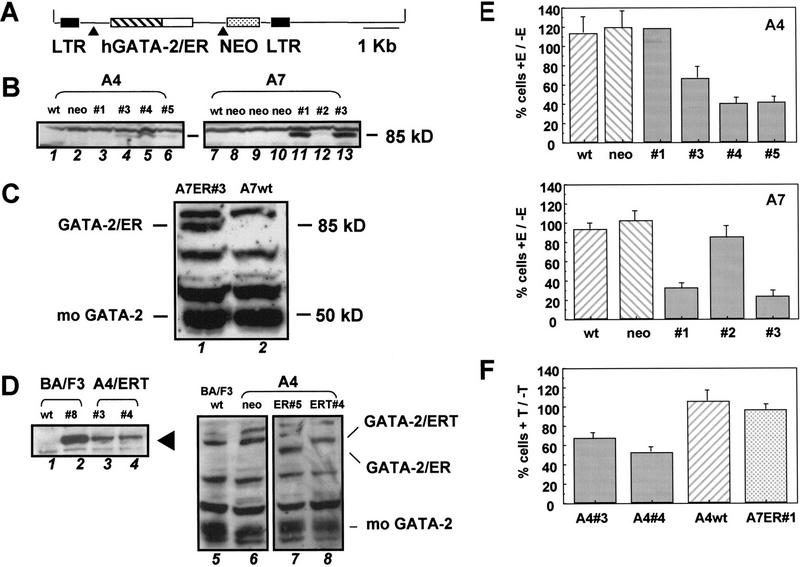
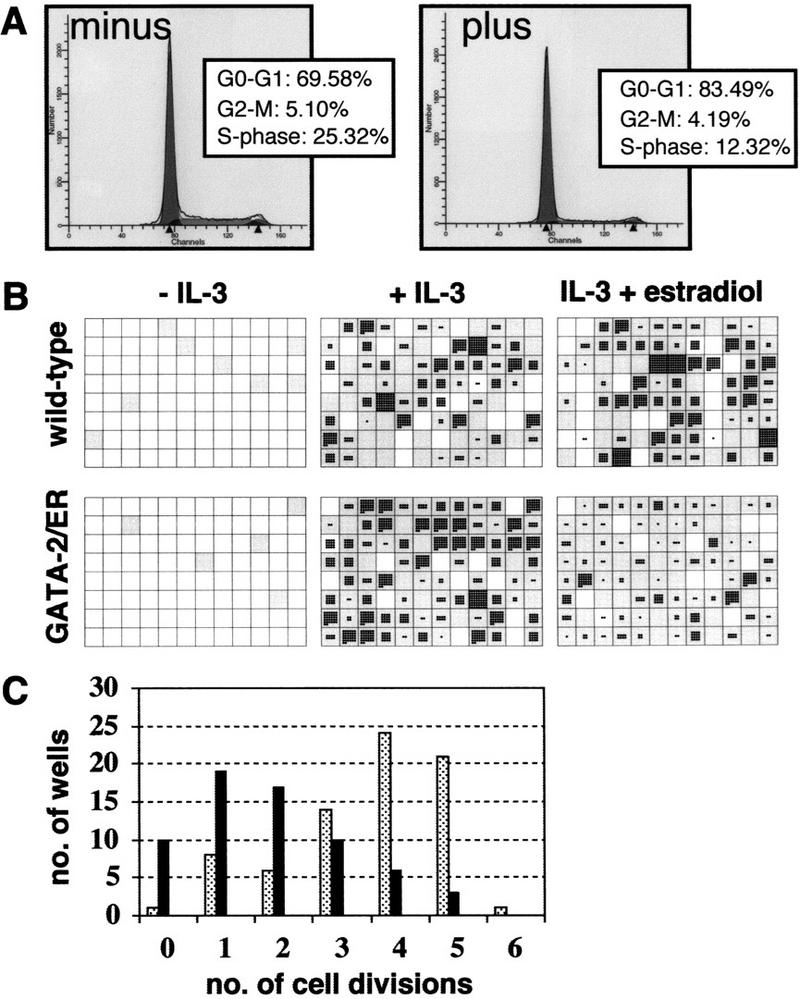
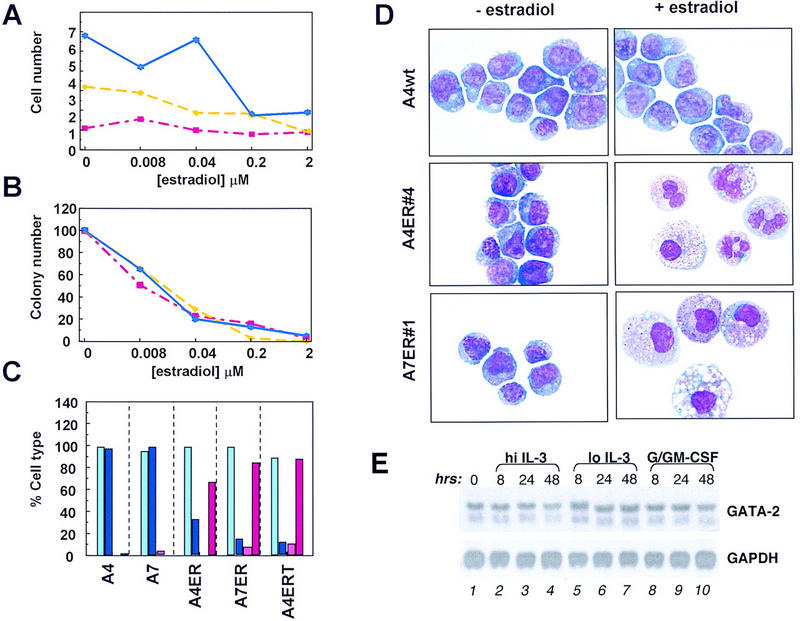
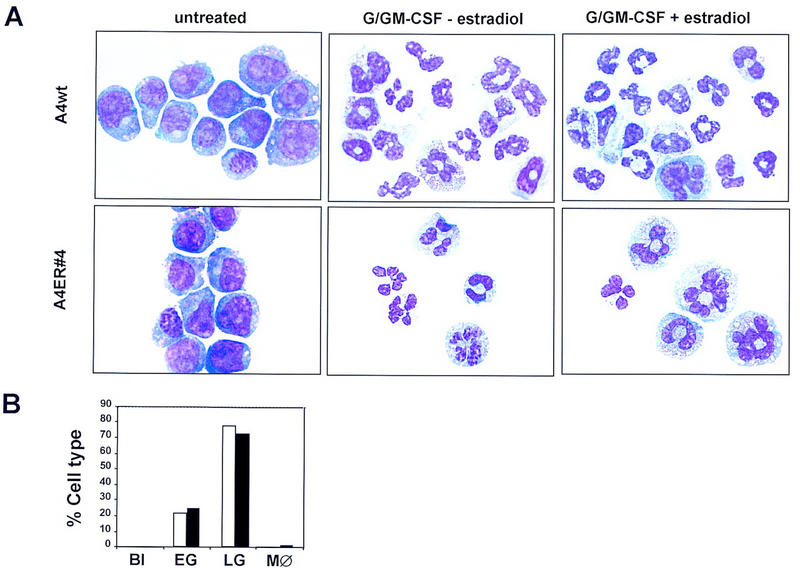
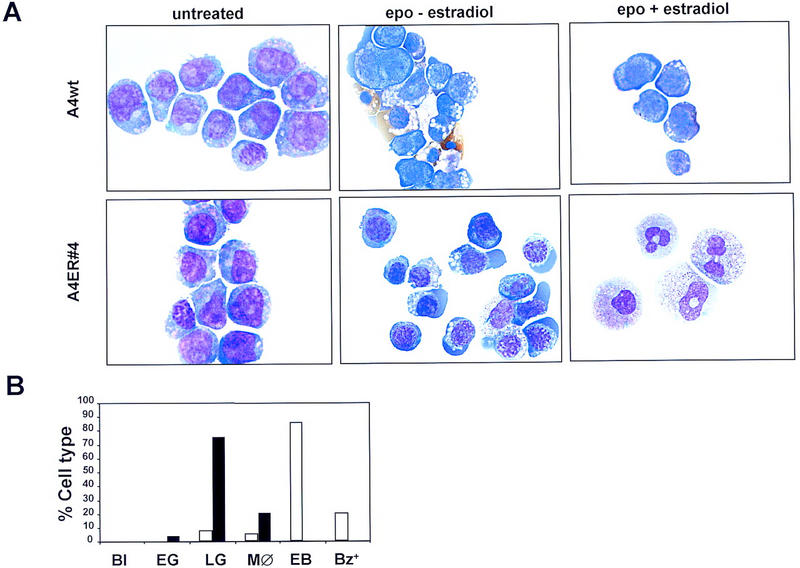
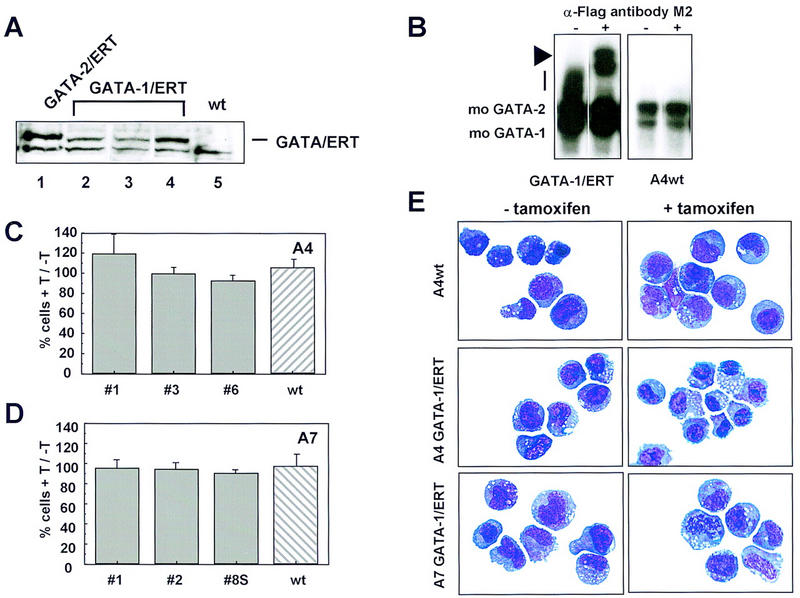
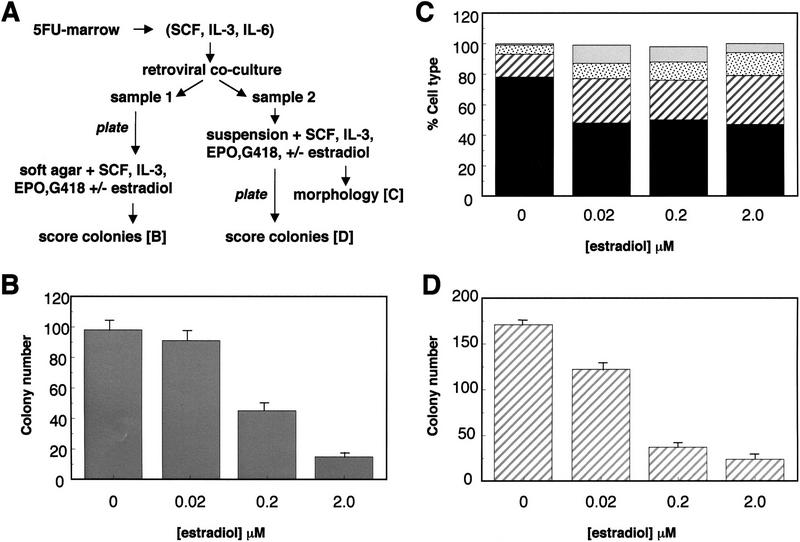
The transcription factor GATA-2 is expressed in hematopoietic stem and progenitor cells and is functionally implicated in their survival and proliferation. We have used estrogen and tamoxifen-inducible forms of GATA-2 to modulate the levels of GATA-2 in the IL-3-dependent multipotential hematopoietic progenitor cell model FDCP mix. Ligand-dependent induction of exogenous GATA-2 activity did not rescue cells deprived of IL-3 from apoptosis. However, induction of GATA-2 activity in cells cultured in IL-3 blocked factor-dependent self-renewal but not factor-dependent survival: Cells undergo cell cycle arrest and cease proliferating but do not apoptose. This was accompanied by differentiation down the monocytic and granulocytic pathways. Differentiation occurred in the presence of IL-3 and did not require addition of exogenous differentiation growth factors such as G-CSF or GM-CSF normally required to induce granulomonocytic differentiation of FDCP-mix cells. Conversely, EPO-dependent erythroid differentiation was inhibited by GATA-2 activation. These biological effects were obtained with levels of exogenous GATA-2 representing less than twofold increases over endogenous GATA-2 levels and were not observed in cells overexpressing GATA-1/ER. Similar effects on proliferation and differentiation were also observed in primary progenitor cells, freshly isolated from murine bone marrow and transduced with a GATA-2/ER-containing retrovirus. Taken together, these data suggest that threshold activities of GATA-2 in hematopoietic progenitor cells are a critical determinant in influencing self-renewal versus differentiation outcomes.
Figures
Similar articles
-
Ectopic expression of a conditional GATA-2/estrogen receptor chimera arrests erythroid differentiation in a hormone-dependent manner.Genes Dev. 1993 Jun;7(6):1097-109. doi: 10.1101/gad.7.6.1097. Genes Dev. 1993. PMID: 8504932
-
GATA-2 and GATA-2/ER display opposing activities in the development and differentiation of blood progenitors.EMBO J. 2002 Jun 17;21(12):3060-9. doi: 10.1093/emboj/cdf301. EMBO J. 2002. PMID: 12065419 Free PMC article.
-
Regulation and function of transcription factor GATA-1 during red blood cell differentiation.Development. 1996 Dec;122(12):3839-50. doi: 10.1242/dev.122.12.3839. Development. 1996. PMID: 9012505
-
Gene expression regulation and domain function of hematopoietic GATA factors.Semin Cell Dev Biol. 2005 Feb;16(1):129-36. doi: 10.1016/j.semcdb.2004.11.001. Epub 2004 Dec 10. Semin Cell Dev Biol. 2005. PMID: 15659347 Review.
-
Roles of hematopoietic transcription factors GATA-1 and GATA-2 in the development of red blood cell lineage.Acta Haematol. 2002;108(4):237-45. doi: 10.1159/000065660. Acta Haematol. 2002. PMID: 12432220 Review.
Cited by
-
Tumor suppressor function of Gata2 in acute promyelocytic leukemia.Blood. 2021 Sep 30;138(13):1148-1161. doi: 10.1182/blood.2021011758. Blood. 2021. PMID: 34125173 Free PMC article.
-
Jun blockade of erythropoiesis: role for repression of GATA-1 by HERP2.Mol Cell Biol. 2004 Sep;24(17):7779-94. doi: 10.1128/MCB.24.17.7779-7794.2004. Mol Cell Biol. 2004. PMID: 15314183 Free PMC article.
-
A single cis element maintains repression of the key developmental regulator Gata2.PLoS Genet. 2010 Sep 9;6(9):e1001103. doi: 10.1371/journal.pgen.1001103. PLoS Genet. 2010. PMID: 20838598 Free PMC article.
-
Transcription factor-mediated lineage switching reveals plasticity in primary committed progenitor cells.EMBO J. 2002 Jul 15;21(14):3770-81. doi: 10.1093/emboj/cdf368. EMBO J. 2002. PMID: 12110589 Free PMC article.
-
Ectopic HOXB4 overcomes the inhibitory effect of tumor necrosis factor-{alpha} on Fanconi anemia hematopoietic stem and progenitor cells.Blood. 2009 May 21;113(21):5111-20. doi: 10.1182/blood-2008-09-180224. Epub 2009 Mar 6. Blood. 2009. PMID: 19270262 Free PMC article.
References
-
- Briegel K, Lim KC, Plank C, Beug H, Engel JD, Zenke M. Ectopic expression of a conditional GATA-2/estrogen receptor chimera arrests erythroid differentiation in a hormone-dependent manner. Genes & Dev. 1993;7:1097–1109. - PubMed
-
- Briegel K, Bartunek P, Stengl G, Lim KC, Beug H, Engel JD, Zenke M. Regulation and function of transcription factor GATA-1 during red blood cell differentiation. Development. 1996;122:3839–3750. - PubMed
-
- Chinnasamy N, Rafferty JA, Hickson I, Lashford LS, Longhurst SJ, Thatcher N, Margison GP, Dexter TM, Fairbairn LJ. Chemoprotective gene transfer II: Multilineage in vivo protection of haemopoiesis against the effects of an antitumour agent by expression of a mutant human O6-alkylguanine-DNA alkyltransferase. Gene Ther. 1998;5:842–847. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials