

Role of the membrane-proximal domain in the initial stages of human immunodeficiency virus type 1 envelope glycoprotein-mediated membrane fusion
- PMID: 10364363
- PMCID: PMC112672
- DOI: 10.1128/JVI.73.7.6089-6092.1999
Role of the membrane-proximal domain in the initial stages of human immunodeficiency virus type 1 envelope glycoprotein-mediated membrane fusion
Abstract
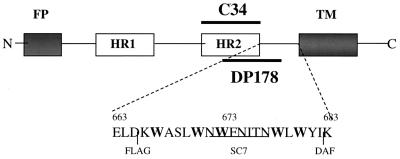
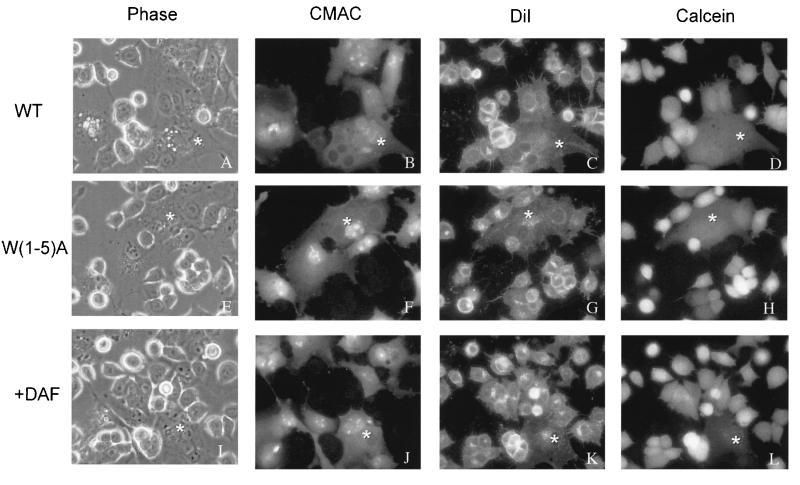
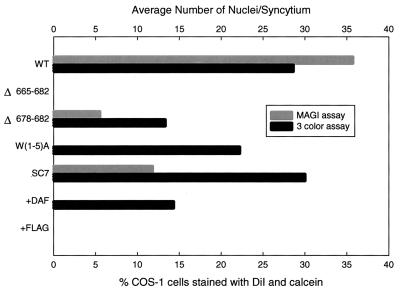
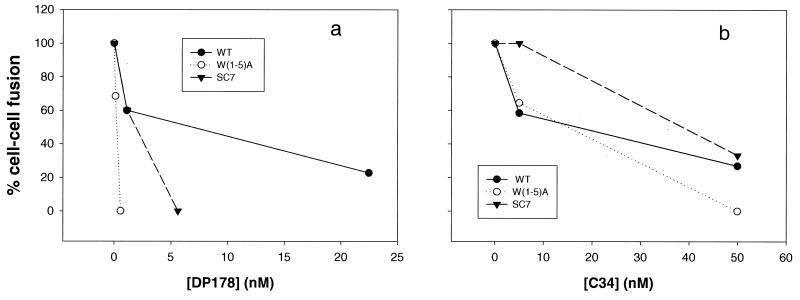
We have examined mutations in the ectodomain of the human immunodeficiency virus type 1 transmembrane glycoprotein gp41 within a region immediately adjacent to the membrane-spanning domain for their effect on the outcome of the fusion cascade. Using the recently developed three-color assay (I. Muñoz-Barroso, S. Durell, K. Sakaguchi, E. Appella, and R. Blumenthal, J. Cell Biol. 140:315-323, 1998), we have assessed the ability of the mutant gp41s to transfer lipid and small solutes from susceptible target cells to the gp120-gp41-expressing cells. The results were compared with the syncytium-inducing capabilities of these gp41 mutants. Two mutant proteins were incapable of mediating both dye transfer and syncytium formation. Two mutant proteins mediated dye transfer but were less effective at inducing syncytium formation than was wild-type gp41. The most interesting mutant proteins were those that were not capable of inducing syncytium formation but still mediated dye transfer, indicating that the fusion cascade was blocked beyond the stage of small fusion pore formation. Fusion mediated by the mutant gp41s was inhibited by the peptides DP178 and C34.
Figures
Similar articles
-
Role of the ectodomain of the gp41 transmembrane envelope protein of human immunodeficiency virus type 1 in late steps of the membrane fusion process.J Virol. 2004 Jan;78(2):811-20. doi: 10.1128/jvi.78.2.811-820.2004. J Virol. 2004. PMID: 14694113 Free PMC article.
-
Target cell-specific determinants of membrane fusion within the human immunodeficiency virus type 1 gp120 third variable region and gp41 amino terminus.J Virol. 1992 Apr;66(4):2389-97. doi: 10.1128/JVI.66.4.2389-2397.1992. J Virol. 1992. PMID: 1548769 Free PMC article.
-
Lack of correlation between soluble CD4-induced shedding of the human immunodeficiency virus type 1 exterior envelope glycoprotein and subsequent membrane fusion events.J Virol. 1992 Sep;66(9):5516-24. doi: 10.1128/JVI.66.9.5516-5524.1992. J Virol. 1992. PMID: 1501286 Free PMC article.
-
Role of hydrophobic residues in the central ectodomain of gp41 in maintaining the association between human immunodeficiency virus type 1 envelope glycoprotein subunits gp120 and gp41.J Virol. 2004 May;78(9):4921-6. doi: 10.1128/jvi.78.9.4921-4926.2004. J Virol. 2004. PMID: 15078976 Free PMC article.
-
CD4 activation of HIV fusion.Int J Cell Cloning. 1992 Nov;10(6):323-32. doi: 10.1002/stem.5530100603. Int J Cell Cloning. 1992. PMID: 1281202 Review.
Cited by
-
Antibodies generated in cats by a lipopeptide reproducing the membrane-proximal external region of the feline immunodeficiency virus transmembrane enhance virus infectivity.Clin Vaccine Immunol. 2007 Aug;14(8):944-51. doi: 10.1128/CVI.00140-07. Epub 2007 Jun 27. Clin Vaccine Immunol. 2007. PMID: 17596431 Free PMC article.
-
Structure of the membrane proximal external region of HIV-1 envelope glycoprotein.Proc Natl Acad Sci U S A. 2018 Sep 18;115(38):E8892-E8899. doi: 10.1073/pnas.1807259115. Epub 2018 Sep 5. Proc Natl Acad Sci U S A. 2018. PMID: 30185554 Free PMC article.
-
Common principles and intermediates of viral protein-mediated fusion: the HIV-1 paradigm.Retrovirology. 2008 Dec 10;5:111. doi: 10.1186/1742-4690-5-111. Retrovirology. 2008. PMID: 19077194 Free PMC article. Review.
-
Role for the terminal clasp of HIV-1 gp41 glycoprotein in the initiation of membrane fusion.J Biol Chem. 2011 Dec 2;286(48):41331-41343. doi: 10.1074/jbc.M111.299826. Epub 2011 Oct 5. J Biol Chem. 2011. PMID: 21976663 Free PMC article.
-
Identification of the LWYIK motif located in the human immunodeficiency virus type 1 transmembrane gp41 protein as a distinct determinant for viral infection.J Virol. 2009 Jan;83(2):870-83. doi: 10.1128/JVI.01088-08. Epub 2008 Nov 5. J Virol. 2009. PMID: 18987155 Free PMC article.
References
-
- Bullough P A, Hughson F M, Skehel J J, Wiley D C. Structure of influenza haemagglutinin at the pH of membrane fusion [see comments] Nature. 1994;371:37–43. - PubMed
-
- Carr C M, Kim P S. A spring-loaded mechanism for the conformational change of influenza hemagglutinin. Cell. 1993;73:823–832. - PubMed
-
- Chan D C, Fass D, Berger J M, Kim P S. Core structure of gp41 from the HIV envelope glycoprotein. Cell. 1997;89:263–273. - PubMed
-
- Fischer C, Schroth-Diez B, Herrmann A, Garten W, Klenk H D. Acylation of the influenza hemagglutinin modulates fusion activity. Virology. 1998;248:284–294. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
