

Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis
- PMID: 10364156
- PMCID: PMC316772
- DOI: 10.1101/gad.13.11.1382
Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis
Abstract
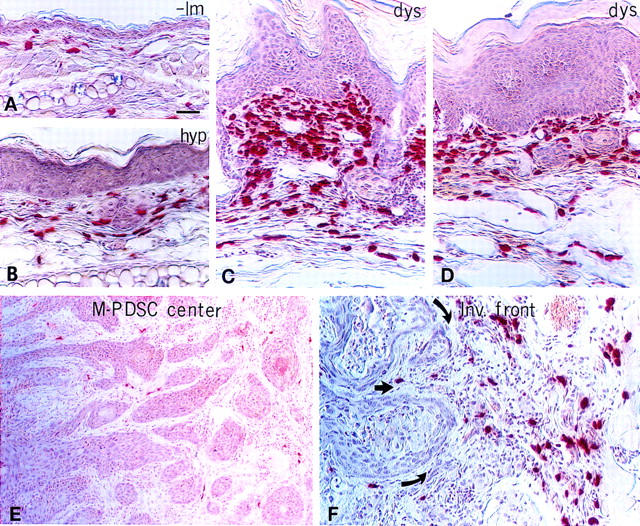
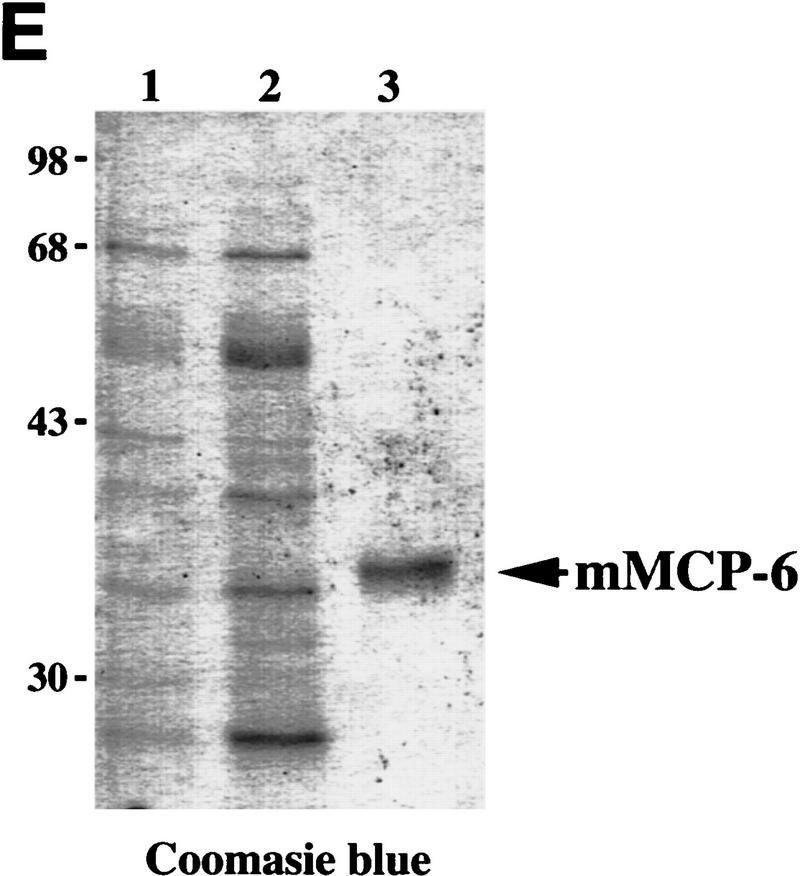
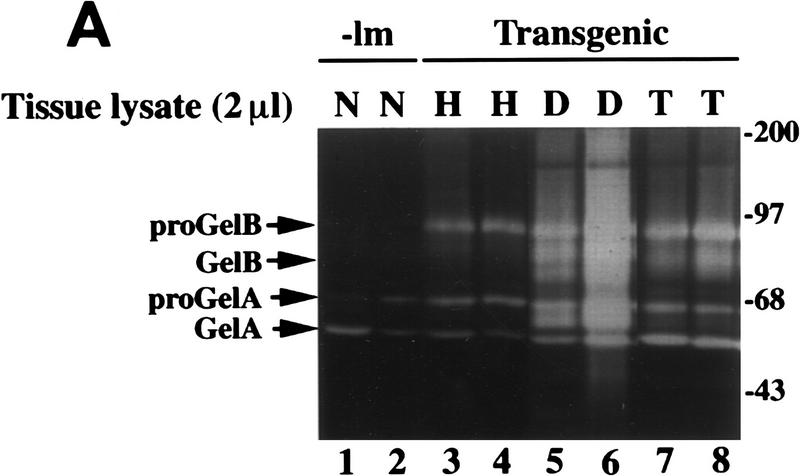
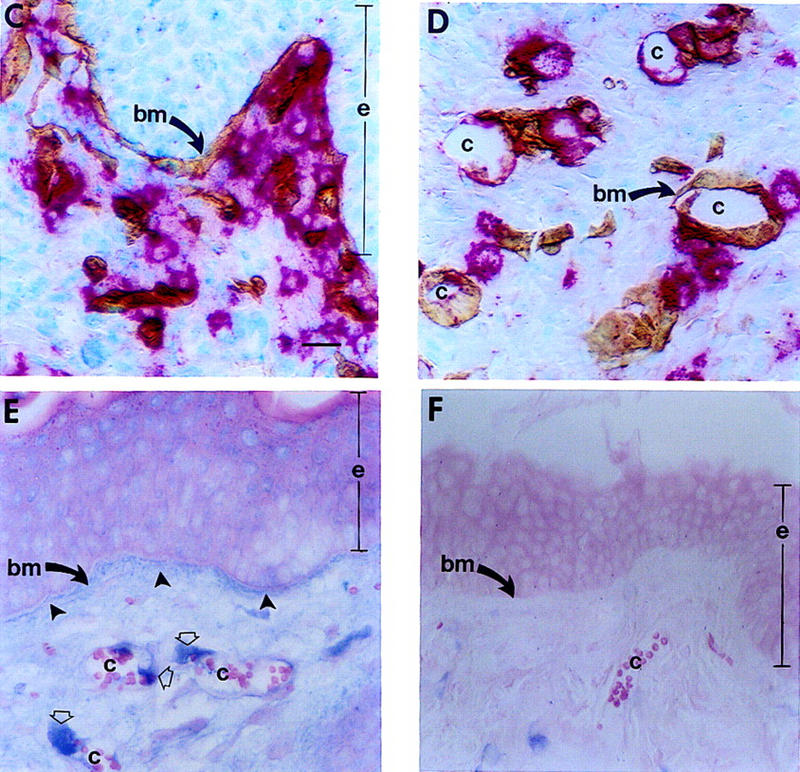
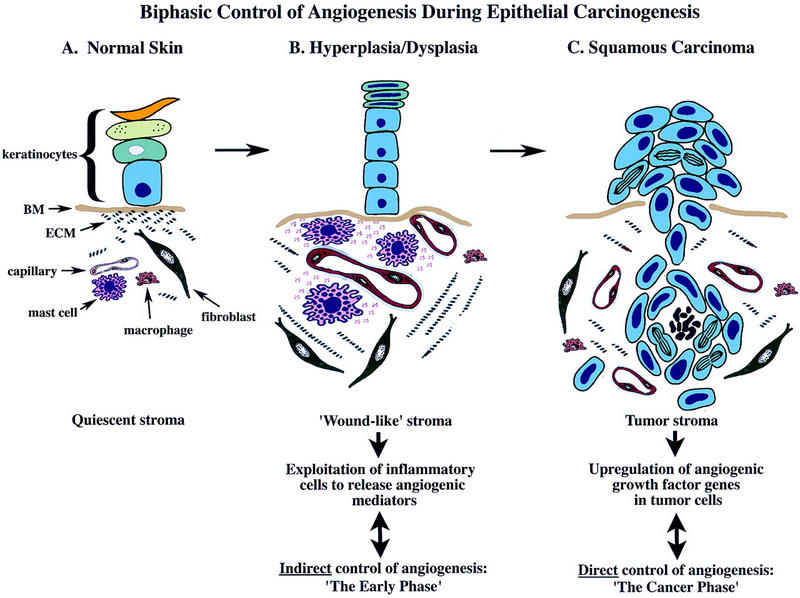
Expression of HPV16 early region genes in basal keratinocytes of transgenic mice elicits a multistage pathway to squamous carcinoma. We report that infiltration by mast cells and activation of the matrix metalloproteinase MMP-9/gelatinase B coincides with the angiogenic switch in premalignant lesions. Mast cells infiltrate hyperplasias, dysplasias, and invasive fronts of carcinomas, but not the core of solid tumors, where they degranulate in close apposition to capillaries and epithelial basement membranes, releasing mast-cell-specific serine proteases MCP-4 (chymase) and MCP-6 (tryptase). MCP-6 is shown to be a mitogen for dermal fibroblasts that proliferate in the reactive stroma, whereas MCP-4 can activate progelatinase B and induce hyperplastic skin to become angiogenic in an in vitro bioassay. Notably, premalignant angiogenesis is abated in a mast-cell-deficient (KITW/KITWWv) HPV16 transgenic mouse. The data indicate that neoplastic progression in this model involves exploitation of an inflammatory response to tissue abnormality. Thus, regulation of angiogenesis during squamous carcinogenesis is biphasic: In hyperplasias, dysplasias, and invading cancer fronts, inflammatory mast cells are conscripted to reorganize stromal architecture and hyperactivate angiogenesis; within the cancer core, upregulation of angiogenesis factors in tumor cells apparently renders them self-sufficient at sustaining neovascularization.
Figures










Similar articles
-
The relationship of tryptase- and chymase-positive mast cells to angiogenesis in stage I non-small cell lung cancer.Eur J Cardiothorac Surg. 2005 Oct;28(4):617-21. doi: 10.1016/j.ejcts.2005.06.020. Eur J Cardiothorac Surg. 2005. PMID: 16125954
-
Direct effects of mast cell proteases, tryptase and chymase, on bronchial epithelial integrity proteins and anti-viral responses.BMC Immunol. 2021 Jun 2;22(1):35. doi: 10.1186/s12865-021-00424-w. BMC Immunol. 2021. PMID: 34078278 Free PMC article.
-
Tissue-specific expression of mast cell granule serine proteinases and their role in inflammation in the lung and gut.Immunology. 2002 Apr;105(4):375-90. doi: 10.1046/j.1365-2567.2002.01375.x. Immunology. 2002. PMID: 11985658 Free PMC article. Review.
-
The role of mast cell tryptase in neoangiogenesis of premalignant and malignant lesions of the uterine cervix.J Histochem Cytochem. 2001 Aug;49(8):1061-2. doi: 10.1177/002215540104900816. J Histochem Cytochem. 2001. PMID: 11457936
-
The Role of Mast Cell Specific Chymases and Tryptases in Tumor Angiogenesis.Biomed Res Int. 2015;2015:142359. doi: 10.1155/2015/142359. Epub 2015 Jun 4. Biomed Res Int. 2015. PMID: 26146612 Free PMC article. Review.
Cited by
-
Different subpopulations of macrophages, neutrophils, mast cells, and fibroblasts are involved in the control of tumor angiogenesis.Front Med (Lausanne). 2024 Oct 8;11:1481609. doi: 10.3389/fmed.2024.1481609. eCollection 2024. Front Med (Lausanne). 2024. PMID: 39440039 Free PMC article. Review.
-
TRIP13: A promising cancer immunotherapy target.Cancer Innov. 2024 Oct 11;3(6):e147. doi: 10.1002/cai2.147. eCollection 2024 Dec. Cancer Innov. 2024. PMID: 39398261 Free PMC article. Review.
-
Milestones in tumor vascularization and its therapeutic targeting.Nat Cancer. 2024 Jun;5(6):827-843. doi: 10.1038/s43018-024-00780-7. Epub 2024 Jun 25. Nat Cancer. 2024. PMID: 38918437 Review.
-
Stabilizing Tumor-Resident Mast Cells Restores T-Cell Infiltration and Sensitizes Sarcomas to PD-L1 Inhibition.Clin Cancer Res. 2024 Jun 3;30(11):2582-2597. doi: 10.1158/1078-0432.CCR-24-0246. Clin Cancer Res. 2024. PMID: 38578281 Free PMC article.
-
The cellular triumvirate: fibroblasts entangled in the crosstalk between cancer cells and immune cells.Front Immunol. 2024 Jan 18;14:1337333. doi: 10.3389/fimmu.2023.1337333. eCollection 2023. Front Immunol. 2024. PMID: 38313431 Free PMC article. Review.
References
-
- Aaltomaa S, Lipponen P, Papinaho S, Kosma V-M. Mast cells in breast cancer. Anticancer Res. 1993;13:785–788. - PubMed
-
- Adler RR, Brenner CA, Werb Z. Expression of extracellular matrix-degrading metalloproteinases and metalloproteinase inhibitors is developmentally regulated during endoderm differentiation of embryonal carcinoma cells. Development. 1990;110:211–220. - PubMed
-
- Arbeit JM, Olson D, Hanahan D. Upregulation of fibroblast growth factors and their receptors during multi-stage epidermal carcinogenesis in K14–HPV16 transgenic mice. Oncogene. 1996;13:1847–1857. - PubMed
-
- Bashkin P, Razin E, Eldor A, Vlodavsky I. Degranulating mast cells secrete an endoglycosidase which degrades heparin sulfate in subendothelial extracellular matrix. Blood. 1990;75:2204–32212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous