

Differential roles for cyclin-dependent kinase inhibitors p21 and p16 in the mechanisms of senescence and differentiation in human fibroblasts
- PMID: 10022898
- PMCID: PMC84004
- DOI: 10.1128/MCB.19.3.2109
Differential roles for cyclin-dependent kinase inhibitors p21 and p16 in the mechanisms of senescence and differentiation in human fibroblasts
Abstract
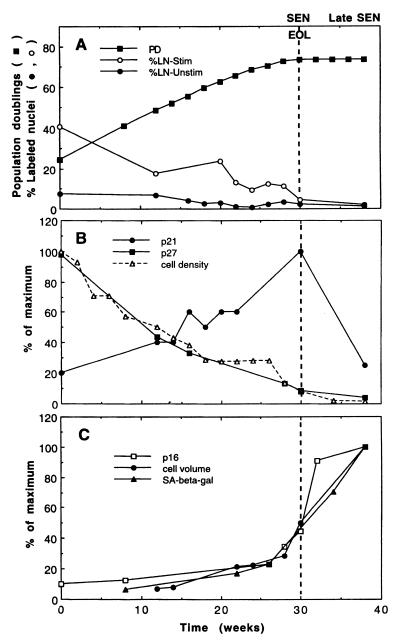
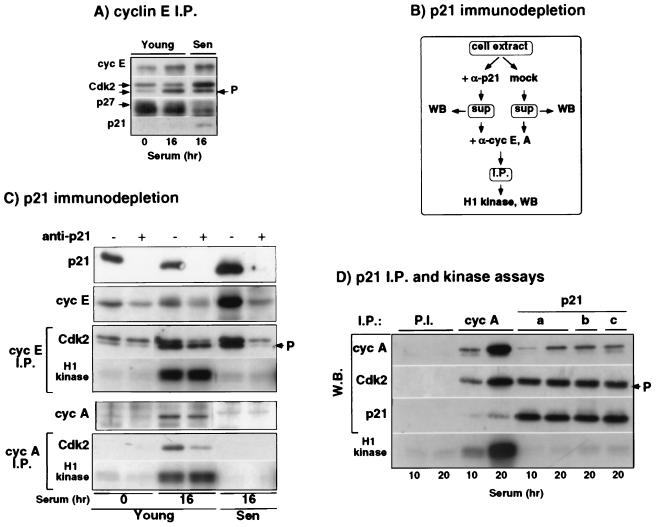
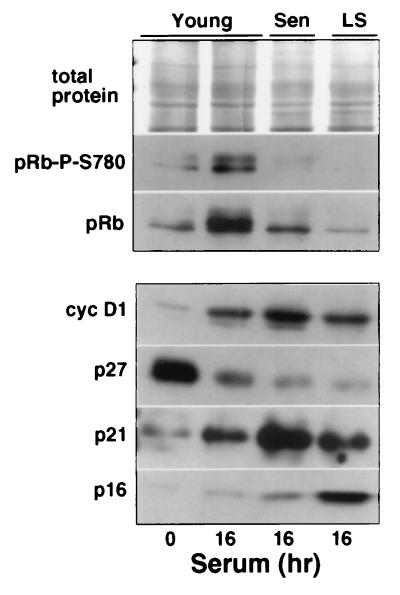
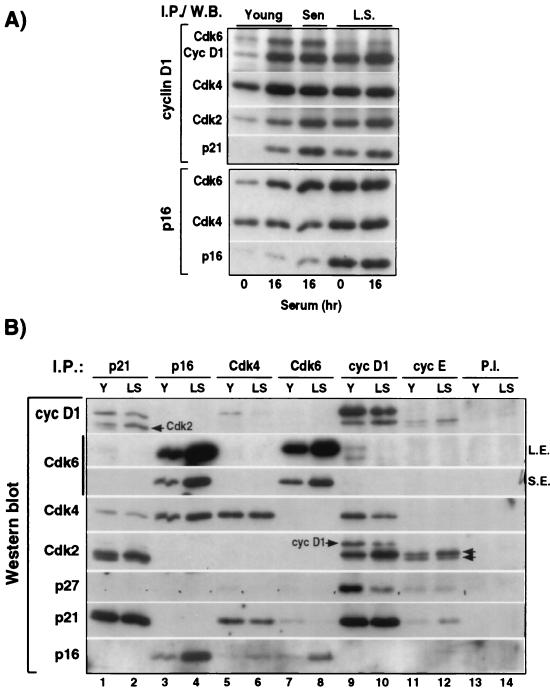
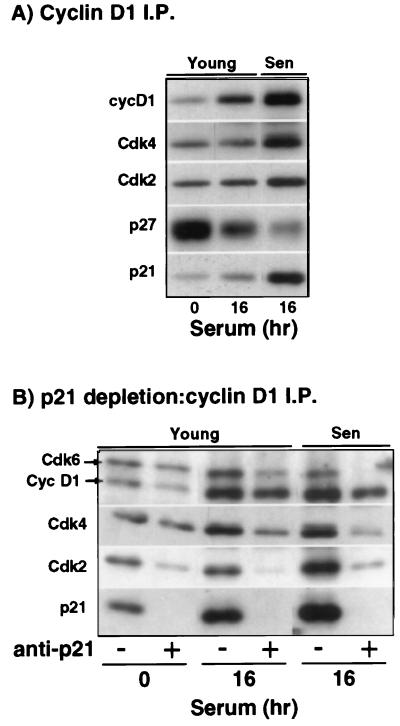
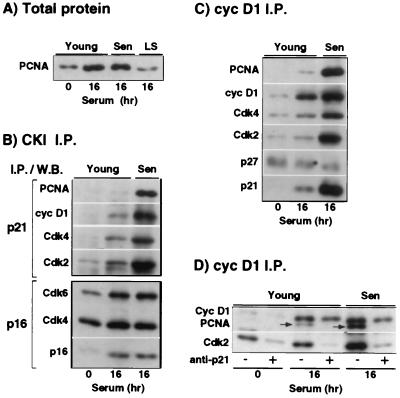
The irreversible G1 arrest in senescent human diploid fibroblasts is probably caused by inactivation of the G1 cyclin-cyclin-dependent kinase (Cdk) complexes responsible for phosphorylation of the retinoblastoma protein (pRb). We show that the Cdk inhibitor p21(Sdi1,Cip1,Waf1), which accumulates progressively in aging cells, binds to and inactivates all cyclin E-Cdk2 complexes in senescent cells, whereas in young cells only p21-free Cdk2 complexes are active. Furthermore, the senescent-cell-cycle arrest occurs prior to the accumulation of the Cdk4-Cdk6 inhibitor p16(Ink4a), suggesting that p21 may be sufficient for this event. Accordingly, cyclin D1-associated phosphorylation of pRb at Ser-780 is lacking even in newly senescent fibroblasts that have a low amount of p16. Instead, the cyclin D1-Cdk4 and cyclin D1-Cdk6 complexes in these cells are associated with an increased amount of p21, suggesting that p21 may be responsible for inactivation of both cyclin E- and cyclin D1-associated kinase activity at the early stage of senescence. Moreover, even in the late stage of senescence when p16 is high, cyclin D1-Cdk4 complexes are persistent, albeit reduced by </=50% compared to young cells. We also provide new evidence that p21 may play a role in inactivation of the DNA replication factor proliferating cell nuclear antigen during early senescence. Finally, because p16 accumulates in parallel with the increases in senescence-associated beta-Gal activity and cell volume that characterize the senescent phenotype, we suggest that p16 upregulation may be part of a differentiation program that is turned on in senescent cells. Since p21 decreases after senescence is achieved, this upregulation of p16 may be essential for maintenance of the senescent-cell-cycle arrest.
Figures
Similar articles
-
Uncoupling between phenotypic senescence and cell cycle arrest in aging p21-deficient fibroblasts.Mol Cell Biol. 2000 Sep;20(18):6741-54. doi: 10.1128/MCB.20.18.6741-6754.2000. Mol Cell Biol. 2000. PMID: 10958672 Free PMC article.
-
Complex mechanisms underlying impaired activation of Cdk4 and Cdk2 in replicative senescence: roles of p16, p21, and cyclin D1.Exp Cell Res. 1999 Dec 15;253(2):503-10. doi: 10.1006/excr.1999.4698. Exp Cell Res. 1999. PMID: 10585273
-
Multifaceted regulation of cell cycle progression by estrogen: regulation of Cdk inhibitors and Cdc25A independent of cyclin D1-Cdk4 function.Mol Cell Biol. 2001 Feb;21(3):794-810. doi: 10.1128/MCB.21.3.794-810.2001. Mol Cell Biol. 2001. PMID: 11154267 Free PMC article.
-
[Molecular mechanisms controlling the cell cycle: fundamental aspects and implications for oncology].Cancer Radiother. 2001 Apr;5(2):109-29. doi: 10.1016/s1278-3218(01)00087-7. Cancer Radiother. 2001. PMID: 11355576 Review. French.
-
Where the cell cycle and histones meet.Genes Dev. 2000 Sep 15;14(18):2265-70. doi: 10.1101/gad.842100. Genes Dev. 2000. PMID: 10995383 Review. No abstract available.
Cited by
-
Role of Cdkn2a in the Emery-Dreifuss Muscular Dystrophy Cardiac Phenotype.Biomolecules. 2021 Apr 6;11(4):538. doi: 10.3390/biom11040538. Biomolecules. 2021. PMID: 33917623 Free PMC article.
-
Chondrocytes From Osteoarthritic and Chondrocalcinosis Cartilage Represent Different Phenotypes.Front Cell Dev Biol. 2021 Apr 26;9:622287. doi: 10.3389/fcell.2021.622287. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33981699 Free PMC article.
-
IGFBP-rP1 induces p21 expression through a p53-independent pathway, leading to cellular senescence of MCF-7 breast cancer cells.J Cancer Res Clin Oncol. 2012 Jun;138(6):1045-55. doi: 10.1007/s00432-012-1153-y. Epub 2012 Mar 6. J Cancer Res Clin Oncol. 2012. PMID: 22392074
-
Iron deprivation in cancer--potential therapeutic implications.Nutrients. 2013 Jul 24;5(8):2836-59. doi: 10.3390/nu5082836. Nutrients. 2013. PMID: 23887041 Free PMC article. Review.
-
Persistent alveolar type 2 dysfunction and lung structural derangement in post-acute COVID-19.medRxiv [Preprint]. 2022 Nov 29:2022.11.28.22282811. doi: 10.1101/2022.11.28.22282811. medRxiv. 2022. PMID: 36482970 Free PMC article. Preprint.
References
-
- Bresnahan W A, Boldogh I, Ma T, Albrecht T, Thompson E A. Cyclin E/Cdk2 activity is controlled by different mechanisms in the G0 and G1 phases of the cell cycle. Cell Growth Differ. 1996;7:1283–1290. - PubMed
-
- Brown J P, Wei W, Sedivy J M. Bypass of senescence after disruption of p21Cip1/Waf1 gene in normal diploid human fibroblasts. Science. 1997;277:831–834. - PubMed
-
- Campisi J. The biology of replicative senescence. Eur J Cancer. 1997;33:703–709. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials