

Hir proteins are required for position-dependent gene silencing in Saccharomyces cerevisiae in the absence of chromatin assembly factor I
- PMID: 9671489
- PMCID: PMC109065
- DOI: 10.1128/MCB.18.8.4793
Hir proteins are required for position-dependent gene silencing in Saccharomyces cerevisiae in the absence of chromatin assembly factor I
Abstract
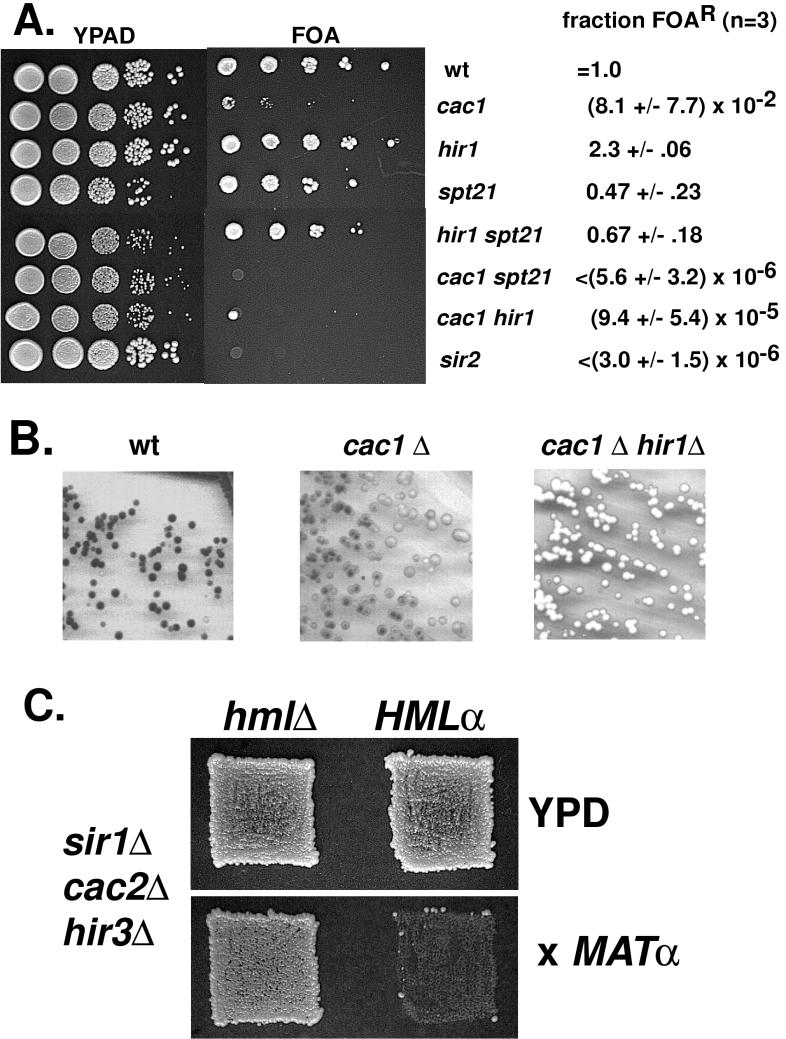
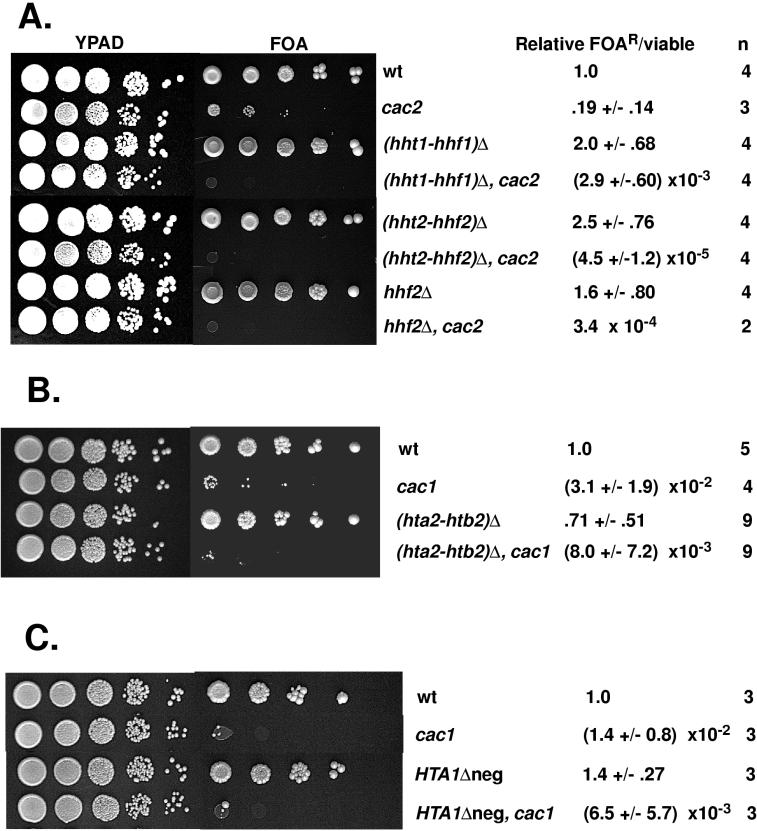
Chromatin assembly factor I (CAF-I) is a three-subunit histone-binding complex conserved from the yeast Saccharomyces cerevisiae to humans. Yeast cells lacking CAF-I (cacDelta mutants) have defects in heterochromatic gene silencing. In this study, we showed that deletion of HIR genes, which regulate histone gene expression, synergistically reduced gene silencing at telomeres and at the HM loci in cacDelta mutants, although hirDelta mutants had no silencing defects when CAF-I was intact. Therefore, Hir proteins are required for an alternative silencing pathway that becomes important in the absence of CAF-I. Because Hir proteins regulate expression of histone genes, we tested the effects of histone gene deletion and overexpression on telomeric silencing and found that alterations in histone H3 and H4 levels or in core histone stoichiometry reduced silencing in cacDelta mutants but not in wild-type cells. We therefore propose that Hir proteins contribute to silencing indirectly via regulation of histone synthesis. However, deletion of combinations of CAC and HIR genes also affected the growth rate and in some cases caused partial temperature sensitivity, suggesting that global aspects of chromosome function may be affected by the loss of members of both gene families.
Figures





Similar articles
-
Chromatin assembly factor I mutants defective for PCNA binding require Asf1/Hir proteins for silencing.Mol Cell Biol. 2002 Jan;22(2):614-25. doi: 10.1128/MCB.22.2.614-625.2002. Mol Cell Biol. 2002. PMID: 11756556 Free PMC article.
-
Yeast histone deposition protein Asf1p requires Hir proteins and PCNA for heterochromatic silencing.Curr Biol. 2001 Apr 3;11(7):463-73. doi: 10.1016/s0960-9822(01)00140-3. Curr Biol. 2001. PMID: 11412995
-
Ultraviolet radiation sensitivity and reduction of telomeric silencing in Saccharomyces cerevisiae cells lacking chromatin assembly factor-I.Genes Dev. 1997 Feb 1;11(3):345-57. doi: 10.1101/gad.11.3.345. Genes Dev. 1997. PMID: 9030687
-
Defects in SPT16 or POB3 (yFACT) in Saccharomyces cerevisiae cause dependence on the Hir/Hpc pathway: polymerase passage may degrade chromatin structure.Genetics. 2002 Dec;162(4):1557-71. doi: 10.1093/genetics/162.4.1557. Genetics. 2002. PMID: 12524332 Free PMC article.
-
Chromatin assembly: the kinetochore connection.Curr Biol. 2002 Apr 2;12(7):R256-8. doi: 10.1016/s0960-9822(02)00786-8. Curr Biol. 2002. PMID: 11937044 Review.
Cited by
-
The C terminus of the histone chaperone Asf1 cross-links to histone H3 in yeast and promotes interaction with histones H3 and H4.Mol Cell Biol. 2013 Feb;33(3):605-21. doi: 10.1128/MCB.01053-12. Epub 2012 Nov 26. Mol Cell Biol. 2013. PMID: 23184661 Free PMC article.
-
Schizosaccharomyces pombe Hat1 (Kat1) is associated with Mis16 and is required for telomeric silencing.Eukaryot Cell. 2012 Sep;11(9):1095-103. doi: 10.1128/EC.00123-12. Epub 2012 Jul 6. Eukaryot Cell. 2012. PMID: 22771823 Free PMC article.
-
Chromatin assembly factor I mutants defective for PCNA binding require Asf1/Hir proteins for silencing.Mol Cell Biol. 2002 Jan;22(2):614-25. doi: 10.1128/MCB.22.2.614-625.2002. Mol Cell Biol. 2002. PMID: 11756556 Free PMC article.
-
The histone shuffle: histone chaperones in an energetic dance.Trends Biochem Sci. 2010 Sep;35(9):476-89. doi: 10.1016/j.tibs.2010.04.001. Epub 2010 May 3. Trends Biochem Sci. 2010. PMID: 20444609 Free PMC article. Review.
-
O-linked N-acetylglucosamine transferase (OGT) interacts with the histone chaperone HIRA complex and regulates nucleosome assembly and cellular senescence.Proc Natl Acad Sci U S A. 2016 Jun 7;113(23):E3213-20. doi: 10.1073/pnas.1600509113. Epub 2016 May 23. Proc Natl Acad Sci U S A. 2016. PMID: 27217568 Free PMC article.
References
-
- Aparicio O M, Billington B L, Gottschling D E. Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell. 1991;66:1279–1287. - PubMed
-
- Chang L, Loranger S S, Mizzen C, Ernst S G, Allis C D, Annunziato A T. Histones in transit: cytosolic histone complexes and diacetylation of H4 during nucleosome assembly in human cells. Biochemistry. 1997;36:469–480. - PubMed
-
- Clark-Adams C D, Norris D, Osley M A, Fassler J S, Winston F. Changes in histone gene dosage alter transcription in yeast. Genes Dev. 1988;2:150–159. - PubMed
-
- Compagnone-Post, P. A., J. Recht, and M. A. Osley. Unpublished data.
-
- Dilworth S M, Black S J, Laskey R A. Two complexes that contain histones are required for nucleosome assembly in vitro: role of nucleoplasmin and N1 in Xenopus egg extracts. Cell. 1987;51:1009–1018. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases