

Identification of positive and negative regulatory regions involved in regulating expression of the human cytomegalovirus UL94 late promoter: role of IE2-86 and cellular p53 in mediating negative regulatory function
- PMID: 9499032
- PMCID: PMC109471
- DOI: 10.1128/JVI.72.3.1814-1825.1998
Identification of positive and negative regulatory regions involved in regulating expression of the human cytomegalovirus UL94 late promoter: role of IE2-86 and cellular p53 in mediating negative regulatory function
Abstract
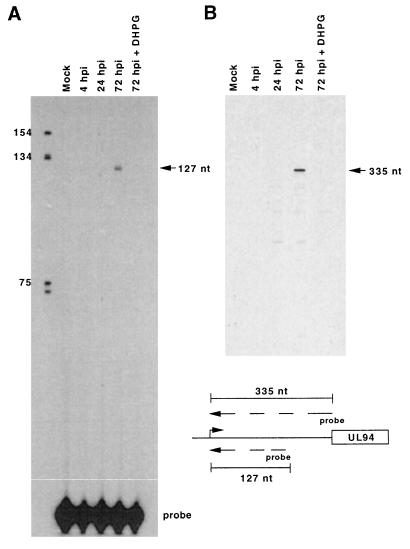
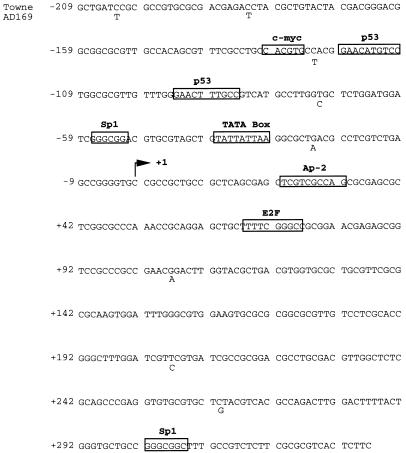
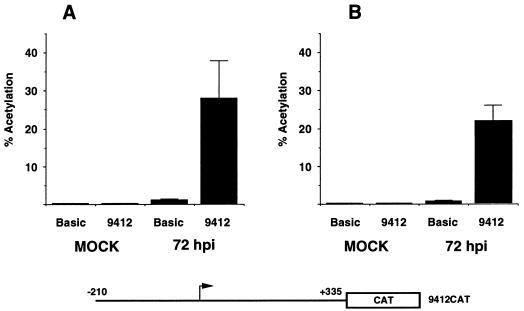
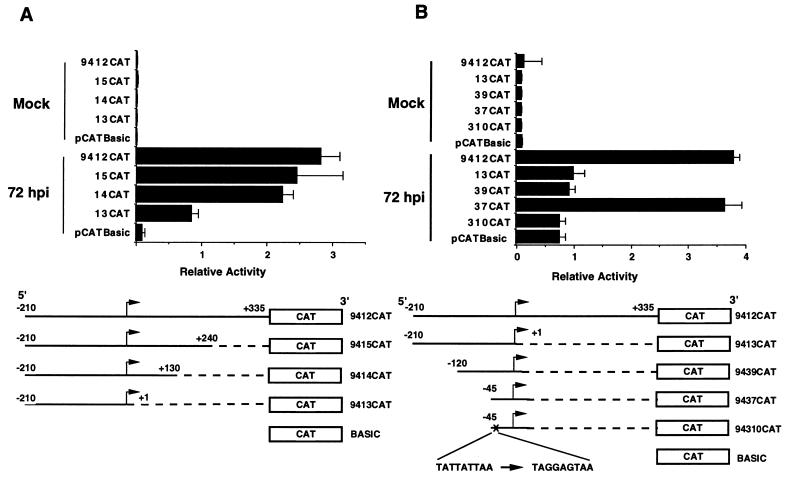
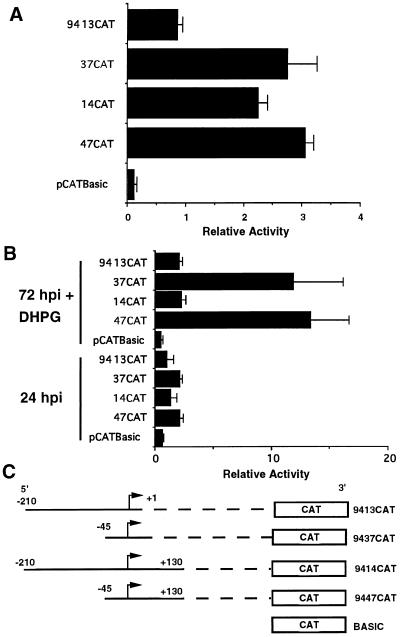
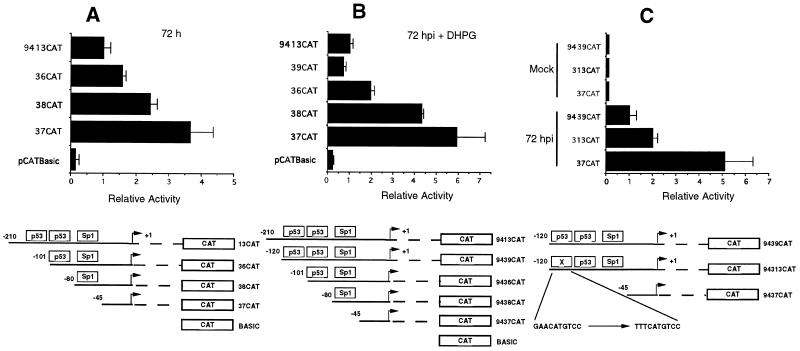
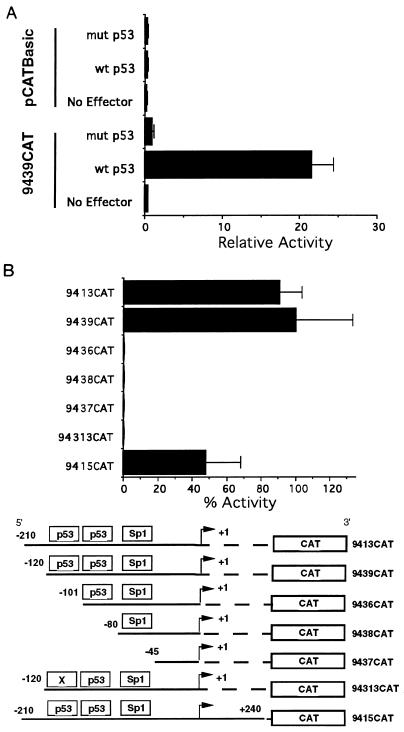
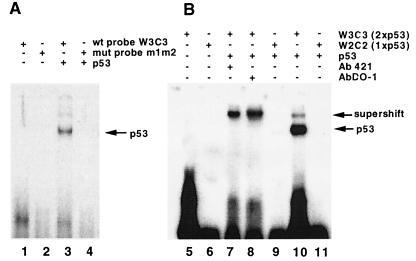
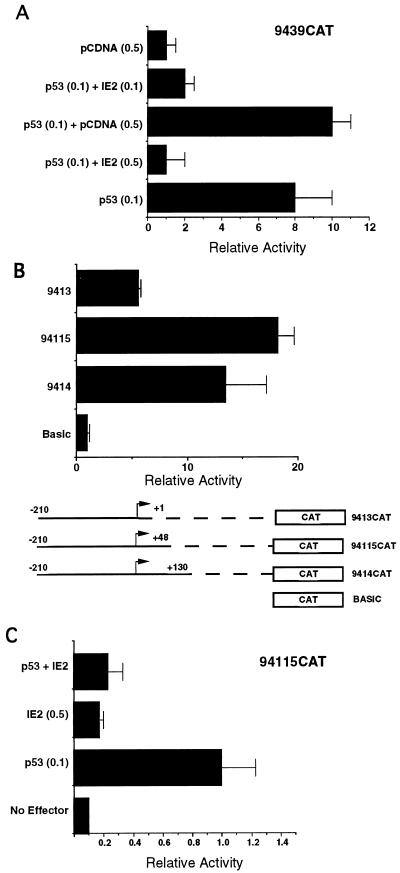
The human cytomegalovirus (HCMV) UL94 gene product is a herpesvirus-common virion protein that is expressed with true late kinetics. To identify the important cis- and trans-acting factors which contribute to UL94 transcriptional regulation, we have cloned, sequenced, and analyzed UL94 promoter function by transient transfection analysis. Transfection of UL94 promoter-reporter gene constructs into permissive human fibroblasts or U373(MG) cells indicated that promoter activity was detected following infection with HCMV. Point mutations within a TATA-like element located upstream of the RNA start site significantly reduced UL94 promoter activity. Deletion mutagenesis of the promoter indicated that a positive regulatory element (PRE) was likely to exist downstream of the UL94 mRNA start site, while a negative regulatory element (NRE) was present upstream of the TATA box. At late times of infection, the PRE appeared to have a dominant effect over the NRE to stimulate maximum levels of UL94 promoter activity, while at earlier times of infection, no activity associated with the PRE could be detected. The NRE, however, appeared to cause constitutive down-regulation of UL94 promoter activity. Binding sites for the cellular p53 protein located within the NRE appeared to contribute to NRE function, and NRE function could be recapitulated in cotransfection assays by concomitant expression of p53 and HCMV IE2-86 protein. Our results suggest a novel mechanism by which the cellular protein p53, which is involved in both transcriptional regulation and progression of cellular DNA synthesis, plays a central role in the regulation of a viral promoter which is not activated prior the onset of viral DNA replication.
Figures
Similar articles
-
Binding of cellular repressor protein or the IE2 protein to a cis-acting negative regulatory element upstream of a human cytomegalovirus early promoter.J Virol. 1995 Dec;69(12):7612-21. doi: 10.1128/JVI.69.12.7612-7621.1995. J Virol. 1995. PMID: 7494269 Free PMC article.
-
A human cytomegalovirus early promoter with upstream negative and positive cis-acting elements: IE2 negates the effect of the negative element, and NF-Y binds to the positive element.J Virol. 1994 Apr;68(4):2108-17. doi: 10.1128/JVI.68.4.2108-2117.1994. J Virol. 1994. PMID: 8138995 Free PMC article.
-
The 72K IE1 and 80K IE2 proteins of human cytomegalovirus independently trans-activate the c-fos, c-myc and hsp70 promoters via basal promoter elements.J Gen Virol. 1992 Sep;73 ( Pt 9):2385-93. doi: 10.1099/0022-1317-73-9-2385. J Gen Virol. 1992. PMID: 1328493
-
Identification of binding sites for the 86-kilodalton IE2 protein of human cytomegalovirus within an IE2-responsive viral early promoter.J Virol. 1994 Jul;68(7):4117-25. doi: 10.1128/JVI.68.7.4117-4125.1994. J Virol. 1994. PMID: 8207790 Free PMC article.
-
Human cytomegalovirus and human herpesvirus 6 genes that transform and transactivate.Clin Microbiol Rev. 1999 Jul;12(3):367-82. doi: 10.1128/CMR.12.3.367. Clin Microbiol Rev. 1999. PMID: 10398670 Free PMC article. Review.
Cited by
-
The p53-microRNA-34a axis regulates cellular entry receptors for tumor-associated human herpes viruses.Med Hypotheses. 2013 Jul;81(1):62-7. doi: 10.1016/j.mehy.2013.04.012. Epub 2013 May 2. Med Hypotheses. 2013. PMID: 23643704 Free PMC article.
-
Human cytomegalovirus transcriptome activity differs during replication in human fibroblast, epithelial and astrocyte cell lines.J Gen Virol. 2012 May;93(Pt 5):1046-1058. doi: 10.1099/vir.0.038083-0. Epub 2012 Jan 18. J Gen Virol. 2012. PMID: 22258857 Free PMC article.
-
Human cytomegalovirus IE1-72 protein interacts with p53 and inhibits p53-dependent transactivation by a mechanism different from that of IE2-86 protein.J Virol. 2009 Dec;83(23):12388-98. doi: 10.1128/JVI.00304-09. Epub 2009 Sep 23. J Virol. 2009. PMID: 19776115 Free PMC article.
-
Potential role for p53 in the permissive life cycle of human cytomegalovirus.J Virol. 2006 Sep;80(17):8390-401. doi: 10.1128/JVI.00505-06. J Virol. 2006. PMID: 16912290 Free PMC article.
-
Transcription of true late (γ2) cytomegalovirus genes requires UL92 function that is conserved among beta- and gammaherpesviruses.J Virol. 2014 Jan;88(1):120-30. doi: 10.1128/JVI.02983-13. Epub 2013 Oct 16. J Virol. 2014. PMID: 24131715 Free PMC article.
References
-
- Alford C A, Britt W J. Cytomegalovirus. In: Fields B N, Knipe D M, Chanock R M, Hirsch M S, Melnick J L, Monath T P, Roizman B, editors. Fields virology. New York, N.Y: Raven Press; 1990. pp. 1981–2010.
-
- Baker S J, Markowitz S, Fearon E R, Willson J K V, Vogelstein B. Supression of human colorectal carcinoma cell growth by wild-type p53. Science. 1990;249:912–915. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
