

A class VI unconventional myosin is associated with a homologue of a microtubule-binding protein, cytoplasmic linker protein-170, in neurons and at the posterior pole of Drosophila embryos
- PMID: 9472041
- PMCID: PMC2141748
- DOI: 10.1083/jcb.140.4.897
A class VI unconventional myosin is associated with a homologue of a microtubule-binding protein, cytoplasmic linker protein-170, in neurons and at the posterior pole of Drosophila embryos
Abstract
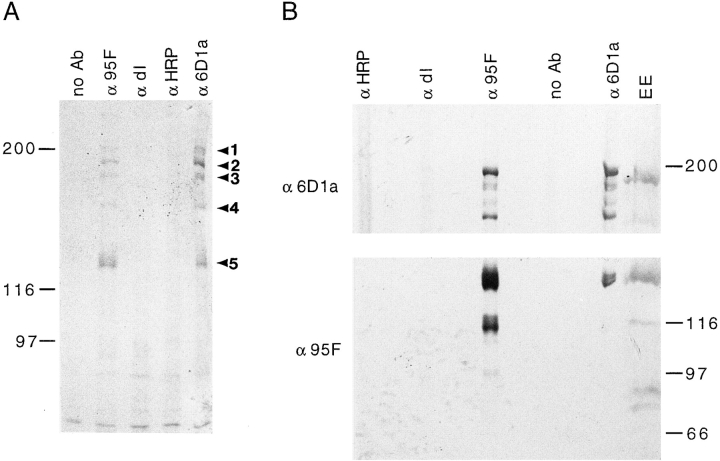
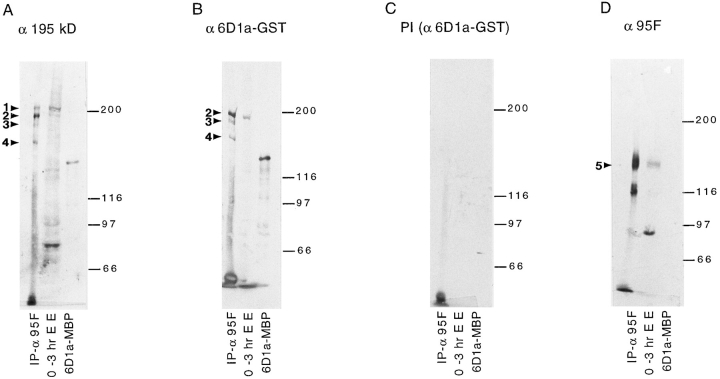
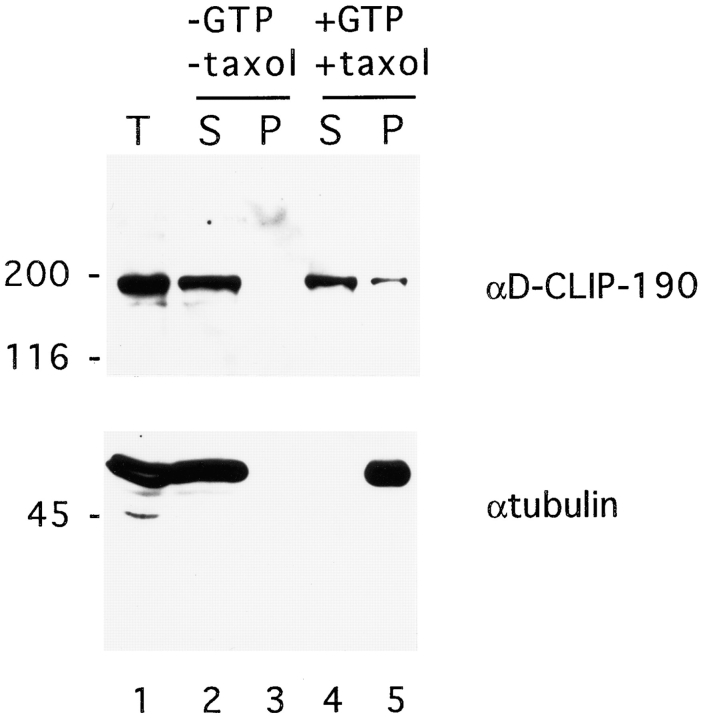
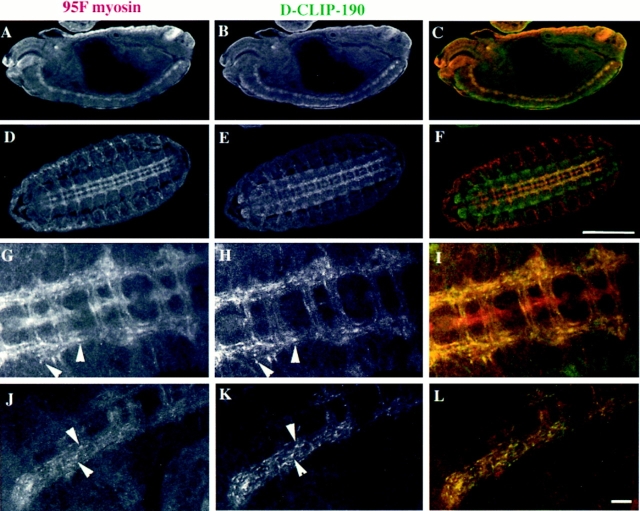
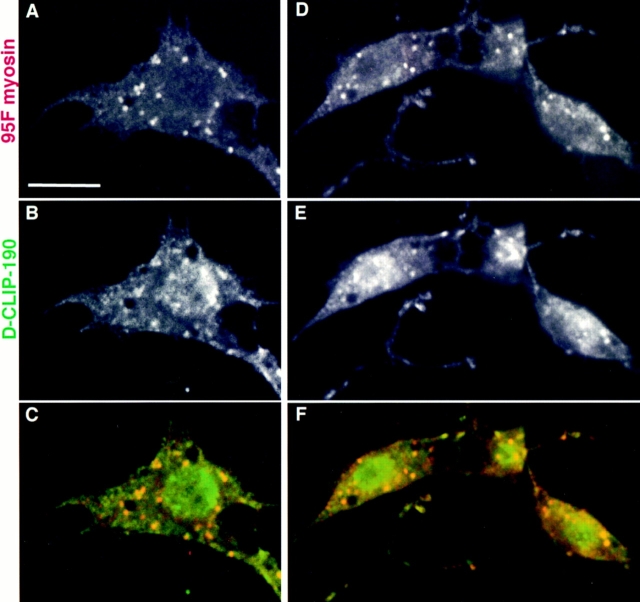
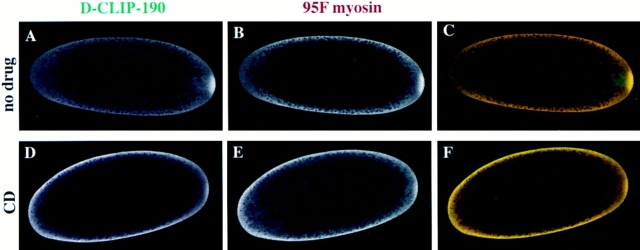
Coordination of cellular organization requires the interaction of the cytoskeletal filament systems. Recently, several lines of investigation have suggested that transport of cellular components along both microtubules and actin filaments is important for cellular organization and function. We report here on molecules that may mediate coordination between the actin and microtubule cytoskeletons. We have identified a 195-kD protein that coimmunoprecipitates with a class VI myosin, Drosophila 95F unconventional myosin. Cloning and sequencing of the gene encoding the 195-kD protein reveals that it is the first homologue identified of cytoplasmic linker protein (CLIP)-170, a protein that links endocytic vesicles to microtubules. We have named this protein D-CLIP-190 (the predicted molecular mass is 189 kD) based on its similarity to CLIP-170 and its ability to cosediment with microtubules. The similarity between D-CLIP-190 and CLIP-170 extends throughout the length of the proteins, and they have a number of predicted sequence and structural features in common. 95F myosin and D-CLIP-190 are coexpressed in a number of tissues during embryogenesis in Drosophila. In the axonal processes of neurons, they are colocalized in the same particulate structures, which resemble vesicles. They are also colocalized at the posterior pole of the early embryo, and this localization is dependent on the actin cytoskeleton. The association of a myosin and a homologue of a microtubule-binding protein in the nervous system and at the posterior pole, where both microtubule and actin-dependent processes are known to be important, leads us to speculate that these two proteins may functionally link the actin and microtubule cytoskeletons.
Figures


Similar articles
-
The 95F unconventional myosin is required for proper organization of the Drosophila syncytial blastoderm.J Cell Biol. 1995 Jun;129(6):1575-88. doi: 10.1083/jcb.129.6.1575. J Cell Biol. 1995. PMID: 7790355 Free PMC article.
-
An unconventional myosin heavy chain gene from Drosophila melanogaster.J Cell Biol. 1992 Nov;119(4):823-34. doi: 10.1083/jcb.119.4.823. J Cell Biol. 1992. PMID: 1429838 Free PMC article.
-
Transport of cytoplasmic particles catalysed by an unconventional myosin in living Drosophila embryos.Nature. 1994 Jun 16;369(6481):560-2. doi: 10.1038/369560a0. Nature. 1994. PMID: 8202156
-
Microtubule-associated proteins as direct crosslinkers of actin filaments and microtubules.IUBMB Life. 2015 Jun;67(6):395-403. doi: 10.1002/iub.1384. Epub 2015 Jun 24. IUBMB Life. 2015. PMID: 26104829 Review.
-
CLIPs and CLASPs and cellular dynamics.Nat Rev Mol Cell Biol. 2005 Jun;6(6):487-98. doi: 10.1038/nrm1664. Nat Rev Mol Cell Biol. 2005. PMID: 15928712 Review.
Cited by
-
Structural determinants for EB1-mediated recruitment of APC and spectraplakins to the microtubule plus end.J Cell Biol. 2005 Feb 14;168(4):587-98. doi: 10.1083/jcb.200410114. Epub 2005 Feb 7. J Cell Biol. 2005. PMID: 15699215 Free PMC article.
-
A dual role for actin and microtubule cytoskeleton in the transport of Golgi units from the nurse cells to the oocyte across ring canals.Mol Biol Cell. 2009 Jan;20(1):556-68. doi: 10.1091/mbc.e08-04-0360. Epub 2008 Nov 12. Mol Biol Cell. 2009. PMID: 19005218 Free PMC article.
-
Drosophila as a genetic and cellular model for studies on axonal growth.Neural Dev. 2007 May 2;2:9. doi: 10.1186/1749-8104-2-9. Neural Dev. 2007. PMID: 17475018 Free PMC article. Review.
-
The localization of myosin VI at the golgi complex and leading edge of fibroblasts and its phosphorylation and recruitment into membrane ruffles of A431 cells after growth factor stimulation.J Cell Biol. 1998 Dec 14;143(6):1535-45. doi: 10.1083/jcb.143.6.1535. J Cell Biol. 1998. PMID: 9852149 Free PMC article.
-
Identification and analysis of the myosin superfamily in Drosophila: a database approach.J Muscle Res Cell Motil. 2000;21(6):491-505. doi: 10.1023/a:1026589626422. J Muscle Res Cell Motil. 2000. PMID: 11206129 Review.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Amero, S.A., T.C. James, and S.C.R. Elgin. 1987. Raising antibodies to protein bands in gels. In Methods in Molecular Biology. Vol. 3. J.M. Walker, editor. Humana Press, Inc., Clifton, NJ. 355–362.
-
- Berg JM. Potential metal-binding domains in nucleic acid binding proteins. Science. 1986;232:485–487. - PubMed
-
- Bilbe G, Delabie J, Bruggen J, Richener H, Asselbergs FAM, Cerletti N, Sorg C, Odink K, Tarcsay L, Wiesendanger W, DeWolf-Peeters C, Shipman R. Restin: a novel intermediate filament-associated protein highly expressed in the Reed-Sternberg cells of Hodgkin's disease. EMBO (Eur Mol Biol Organ) J. 1992;11:2103–2113. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
