

Enhancement of hepatitis B virus infection by noninfectious subviral particles
- PMID: 9445049
- PMCID: PMC124627
- DOI: 10.1128/JVI.72.2.1462-1468.1998
Enhancement of hepatitis B virus infection by noninfectious subviral particles
Abstract
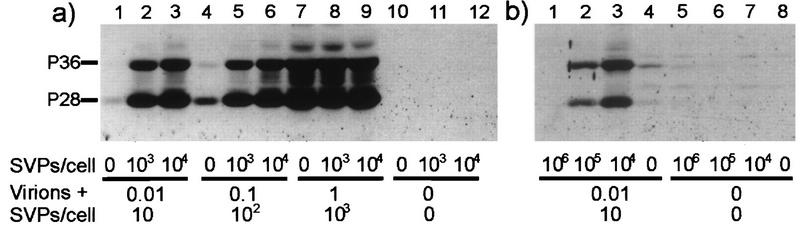
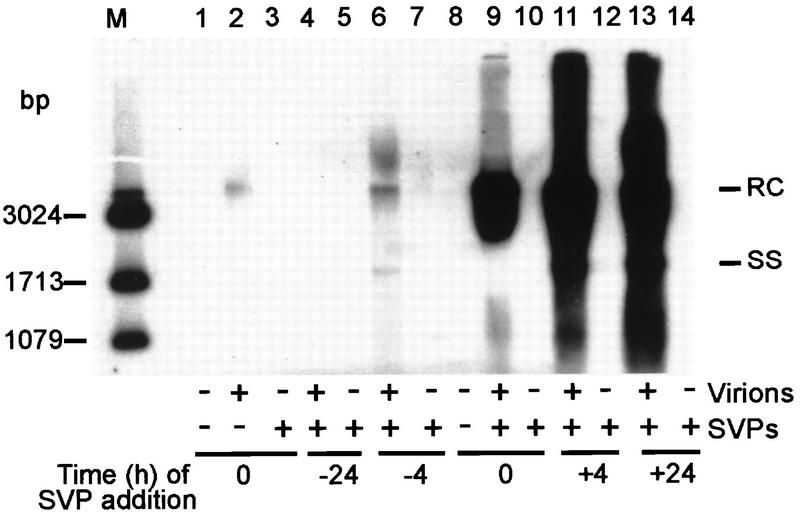
The biological function of the huge excess of subviral particles over virions in hepatitis B virus infections is unknown. Using the duck hepatitis B virus as a model, we unexpectedly found that subviral particles strongly enhance intracellular viral replication and gene expression. This effect is dependent on the multiplicity of infection, the ratio of virions over subviral particles, and the time point of addition of subviral particles. Most importantly, we show that the pre-S protein of the subviral particles triggers enhancement and requires the presence of the binding regions for putative cell-encoded virus receptor proteins. These data suggest that enhancement is due either to the recently described transactivation function of the pre-S protein or to signalling pathways which become activated upon binding of subviral particles to cellular receptors. The findings are of clinical importance, since they imply that infectivity of sera containing hepadnaviruses depends not only on the amount of infectious virions but also decisively on the number of particles devoid of nucleic acids. A similarly dramatic enhancing effect of noninfectious particles in other virus infections is well conceivable.
Figures



Similar articles
-
Common and Distinct Capsid and Surface Protein Requirements for Secretion of Complete and Genome-Free Hepatitis B Virions.J Virol. 2018 Jun 29;92(14):e00272-18. doi: 10.1128/JVI.00272-18. Print 2018 Jul 15. J Virol. 2018. PMID: 29743374 Free PMC article.
-
Hepatitis B Virus-Encoded MicroRNA Controls Viral Replication.J Virol. 2017 Apr 28;91(10):e01919-16. doi: 10.1128/JVI.01919-16. Print 2017 May 15. J Virol. 2017. PMID: 28148795 Free PMC article.
-
Complete and Incomplete Hepatitis B Virus Particles: Formation, Function, and Application.Viruses. 2017 Mar 21;9(3):56. doi: 10.3390/v9030056. Viruses. 2017. PMID: 28335554 Free PMC article. Review.
-
Hepadnavirus infection requires interaction between the viral pre-S domain and a specific hepatocellular receptor.J Virol. 1993 Dec;67(12):7414-22. doi: 10.1128/JVI.67.12.7414-7422.1993. J Virol. 1993. PMID: 8230462 Free PMC article.
-
Hepatitis B virus: The challenge of an ancient virus with multiple faces and a remarkable replication strategy.Antiviral Res. 2018 Oct;158:34-44. doi: 10.1016/j.antiviral.2018.07.019. Epub 2018 Jul 29. Antiviral Res. 2018. PMID: 30059722 Review.
Cited by
-
HBV infection of cell culture: evidence for multivalent and cooperative attachment.EMBO J. 2001 Aug 15;20(16):4443-53. doi: 10.1093/emboj/20.16.4443. EMBO J. 2001. PMID: 11500372 Free PMC article.
-
Carboxypeptidase D (gp180), a Golgi-resident protein, functions in the attachment and entry of avian hepatitis B viruses.J Virol. 1998 Oct;72(10):8098-104. doi: 10.1128/JVI.72.10.8098-8104.1998. J Virol. 1998. PMID: 9733850 Free PMC article.
-
A short note on HBV infection.Bioinformation. 2020 Jul 31;16(7):505-508. doi: 10.6026/97320630016505. eCollection 2020. Bioinformation. 2020. PMID: 32994674 Free PMC article.
-
Properties of subviral particles of hepatitis B virus.J Virol. 2008 Aug;82(16):7812-7. doi: 10.1128/JVI.00561-08. Epub 2008 Jun 4. J Virol. 2008. PMID: 18524834 Free PMC article.
-
Entry of duck hepatitis B virus into primary duck liver and kidney cells after discovery of a fusogenic region within the large surface protein.J Virol. 2007 May;81(10):5014-23. doi: 10.1128/JVI.02290-06. Epub 2007 Mar 14. J Virol. 2007. PMID: 17360753 Free PMC article.
References
-
- Bolander F F., Jr Regulation of the mouse mammary tumor virus receptor by phosphorylation and internalization in mammary epithelial cells. J Cell Physiol. 1994;161:124–128. - PubMed
-
- Bouillie S, Barel M, Drane P, Cassinat B, Balbo M, Holers V M, Frade R. Epstein-Barr virus/C3d receptor (CR2, CD21) activated by its extracellular ligands regulates pp105 phosphorylation through two distinct pathways. Eur J Immunol. 1995;25:2661–2667. - PubMed
-
- Bruns M, Kratzberg T, Zeller W, Lehmann-Grube F. Mode of replication of lymphocytic choriomeningitis virus in persistently infected cultivated mouse L cells. Virology. 1990;177:615–624. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
