

Pex19p, a farnesylated protein essential for peroxisome biogenesis
- PMID: 9418908
- PMCID: PMC121529
- DOI: 10.1128/MCB.18.1.616
Pex19p, a farnesylated protein essential for peroxisome biogenesis
Abstract
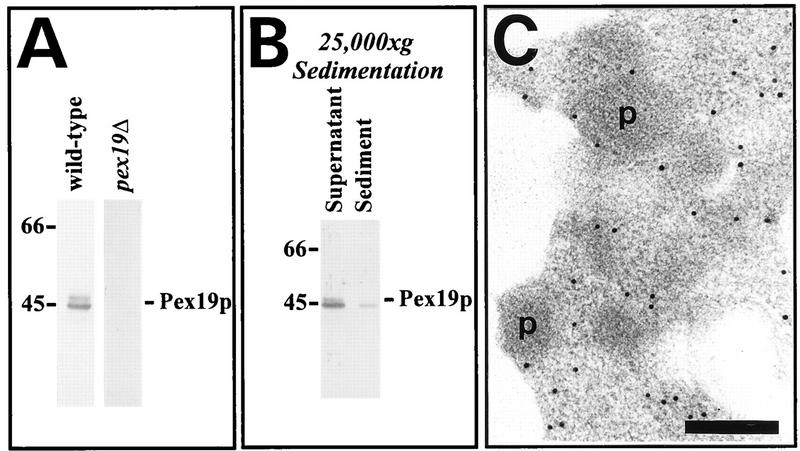
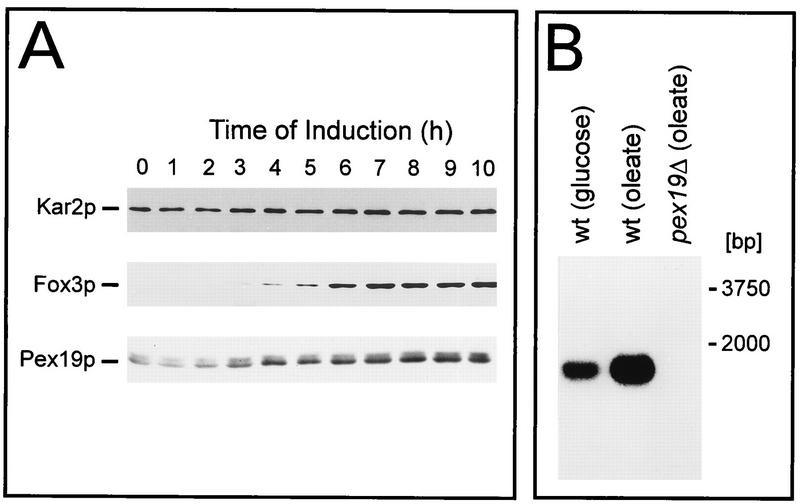
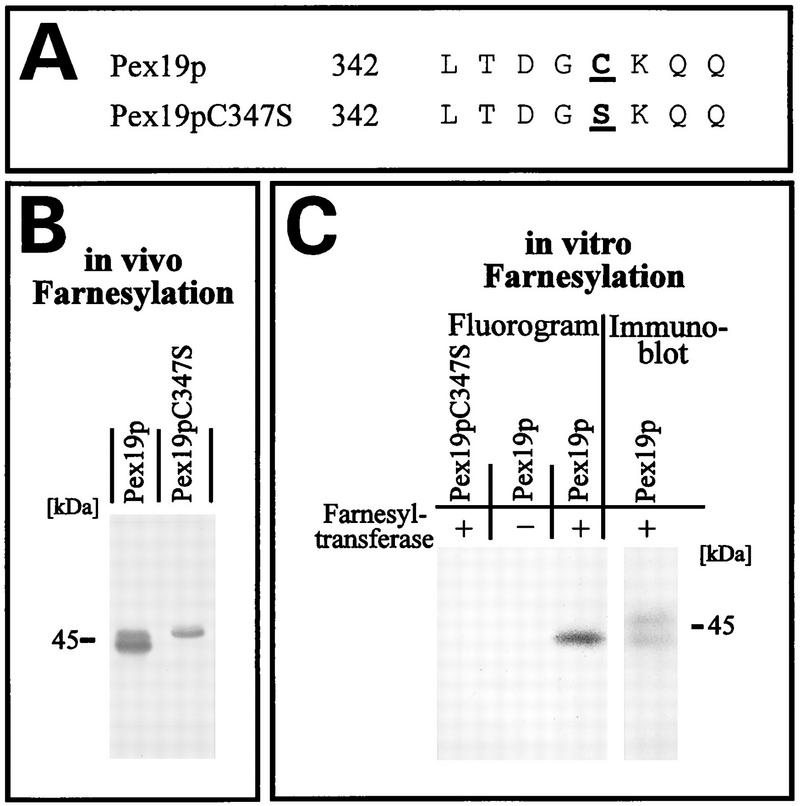
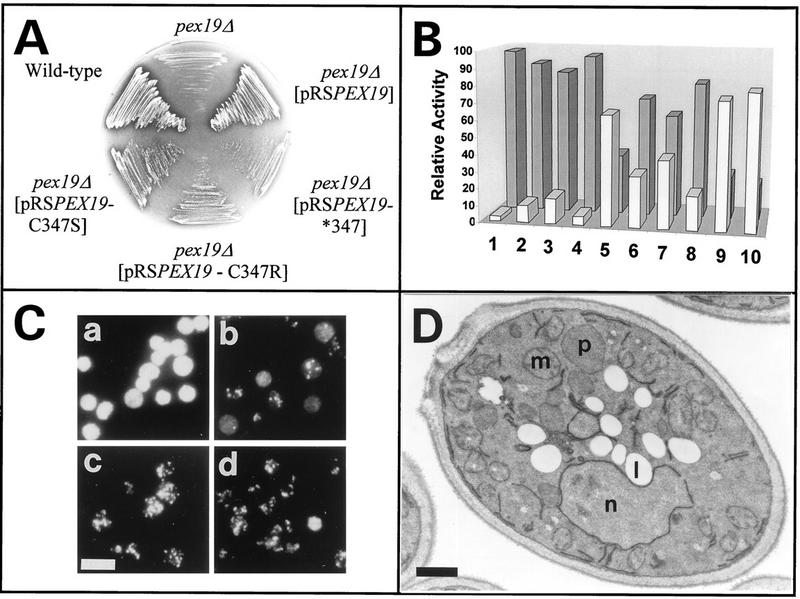
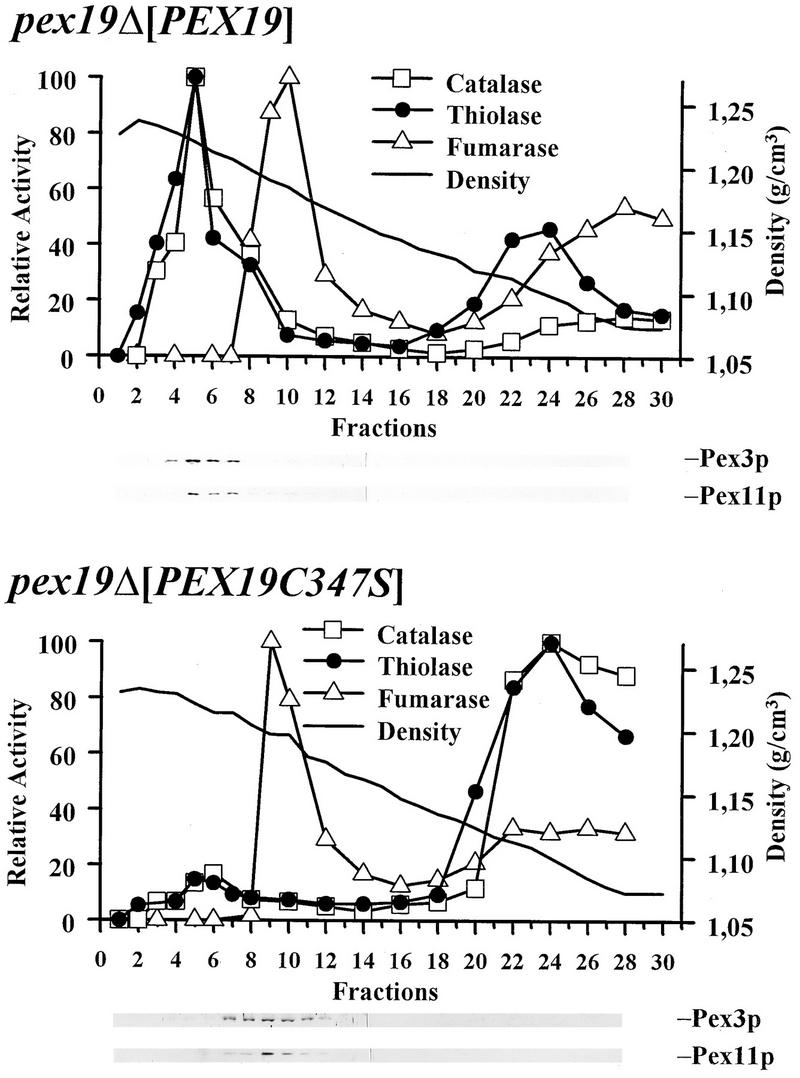
We report the identification and molecular characterization of Pex19p, an oleic acid-inducible, farnesylated protein of 39.7 kDa that is essential for peroxisome biogenesis in Saccharomyces cerevisiae. Cells lacking Pex19p are characterized by the absence of morphologically detectable peroxisomes and mislocalization of peroxisomal matrix proteins to the cytosol. The human HK33 gene product was identified as the putative human ortholog of Pex19p. Evidence is provided that farnesylation of Pex19p takes place at the cysteine of the C-terminal CKQQ amino acid sequence. Farnesylation of Pex19p was shown to be essential for the proper function of the protein in peroxisome biogenesis. Pex19p was shown to interact with Pex3p in vivo, and this interaction required farnesylation of Pex19p.
Figures



Similar articles
-
Pex19p interacts with Pex3p and Pex10p and is essential for peroxisome biogenesis in Pichia pastoris.Mol Biol Cell. 1999 Jun;10(6):1745-61. doi: 10.1091/mbc.10.6.1745. Mol Biol Cell. 1999. PMID: 10359594 Free PMC article.
-
Farnesylation of Pex19p is not essential for peroxisome biogenesis in yeast and mammalian cells.Cell Mol Life Sci. 2006 Jul;63(14):1686-99. doi: 10.1007/s00018-006-6110-y. Cell Mol Life Sci. 2006. PMID: 16791427 Free PMC article.
-
Farnesylation of pex19p is required for its structural integrity and function in peroxisome biogenesis.J Biol Chem. 2009 Jul 31;284(31):20885-96. doi: 10.1074/jbc.M109.016584. Epub 2009 May 18. J Biol Chem. 2009. PMID: 19451657 Free PMC article.
-
Peroxisome biogenesis disorders: molecular basis for impaired peroxisomal membrane assembly: in metabolic functions and biogenesis of peroxisomes in health and disease.Biochim Biophys Acta. 2012 Sep;1822(9):1337-42. doi: 10.1016/j.bbadis.2012.06.004. Epub 2012 Jun 13. Biochim Biophys Acta. 2012. PMID: 22705440 Review.
-
Import of peroxisomal membrane proteins: the interplay of Pex3p- and Pex19p-mediated interactions.Biochim Biophys Acta. 2006 Dec;1763(12):1639-46. doi: 10.1016/j.bbamcr.2006.09.030. Epub 2006 Sep 26. Biochim Biophys Acta. 2006. PMID: 17069900 Review.
Cited by
-
The role of the endoplasmic reticulum in peroxisome biogenesis.Cold Spring Harb Perspect Biol. 2013 May 1;5(5):a013243. doi: 10.1101/cshperspect.a013243. Cold Spring Harb Perspect Biol. 2013. PMID: 23637287 Free PMC article. Review.
-
Elevated Delta-6 desaturase (FADS2) gene expression in the prefrontal cortex of patients with bipolar disorder.J Psychiatr Res. 2011 Feb;45(2):269-72. doi: 10.1016/j.jpsychires.2010.06.010. Epub 2010 Jul 7. J Psychiatr Res. 2011. PMID: 20615514 Free PMC article.
-
Proteolytic processing of certain CaaX motifs can occur in the absence of the Rce1p and Ste24p CaaX proteases.Yeast. 2009 Aug;26(8):451-63. doi: 10.1002/yea.1678. Yeast. 2009. PMID: 19504624 Free PMC article.
-
Analysis of prelamin A biogenesis reveals the nucleus to be a CaaX processing compartment.Mol Biol Cell. 2008 Dec;19(12):5398-408. doi: 10.1091/mbc.e08-07-0704. Epub 2008 Oct 15. Mol Biol Cell. 2008. PMID: 18923140 Free PMC article.
-
The Pex3-Inp1 complex tethers yeast peroxisomes to the plasma membrane.J Cell Biol. 2020 Oct 5;219(10):e201906021. doi: 10.1083/jcb.201906021. J Cell Biol. 2020. PMID: 32970792 Free PMC article.
References
-
- Albertini M, Rehling P, Erdmann R, Girzalsky W, Kiel J A K W, Veenhuis M, Kunau W-H. Pex14p, a peroxisomal membrane protein binding both receptors of the two PTS-dependent import pathways. Cell. 1997;89:1–20. - PubMed
-
- Ausubel F J, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: Greene Publishing Associates and Wiley-Interscience; 1989.
-
- Baerends R J S, Rasmussen S W, Hilbrands R E, van der Heide M, Faber K N, Reuvekamp P T W, Kiel J A K W, Cregg J M, van der Klei I J, Veenhuis M. The Hansenula polymorpha PER9 gene encodes a peroxisomal membrane protein essential for peroxisome assembly and integrity. J Biol Chem. 1996;271:8887–8894. - PubMed
-
- Balch W E. Low molecular weight GTP-binding proteins (LMGPs) involved in vesicular transport: binary switches or biological transducers? Trends Biochem Sci. 1990;15:469–472. - PubMed
-
- Baumgart E. Morphology of peroxisomes in light- and electron microscopy. In: Bugaut M, Latruffe N, editors. Peroxisomes: biochemistry, molecular biology and genetic diseases. Heidelberg, Germany: Springer-Verlag; 1994. pp. 37–57.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases