

Survival of mature CD4 T lymphocytes is dependent on major histocompatibility complex class II-expressing dendritic cells
- PMID: 9334361
- PMCID: PMC2199085
- DOI: 10.1084/jem.186.8.1223
Survival of mature CD4 T lymphocytes is dependent on major histocompatibility complex class II-expressing dendritic cells
Abstract
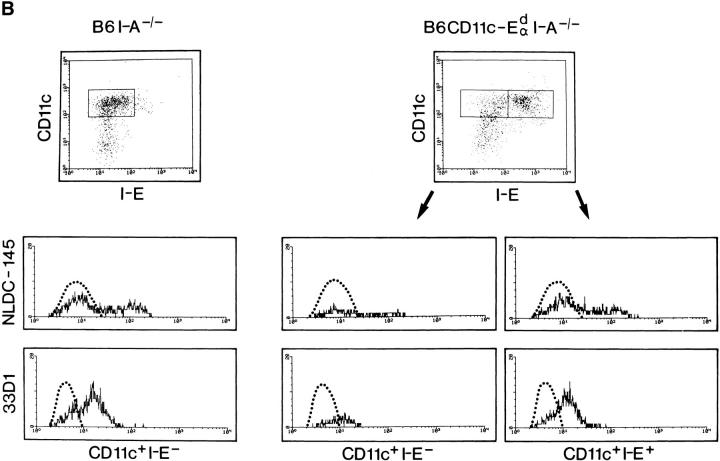
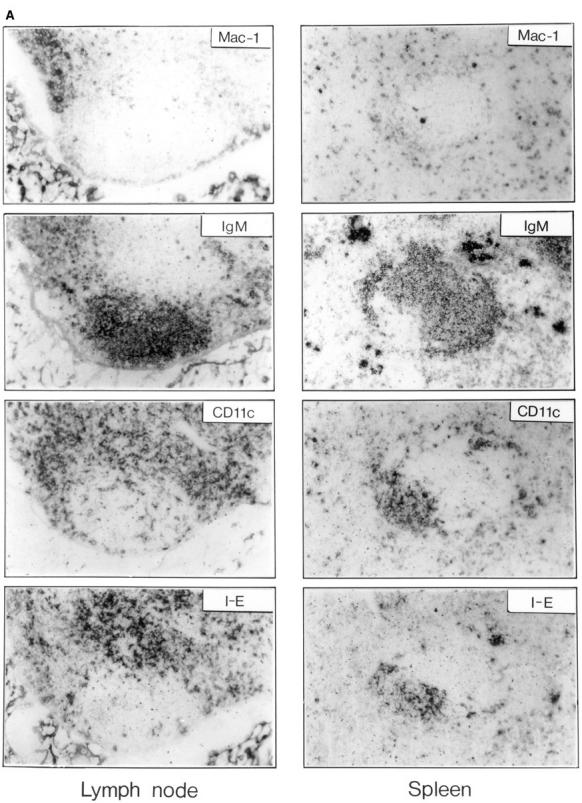
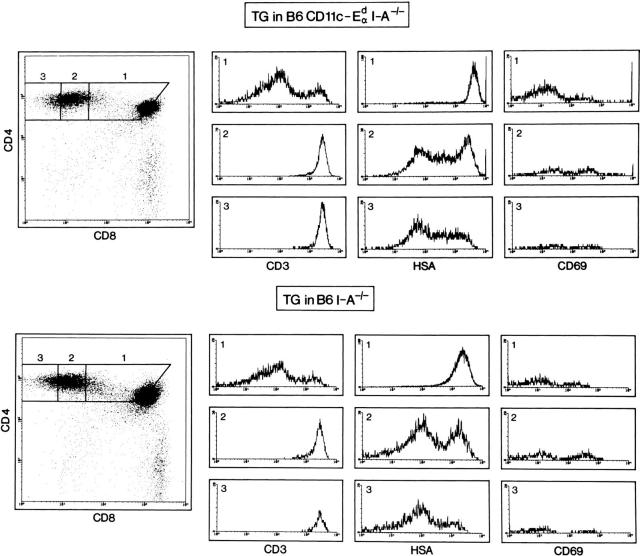
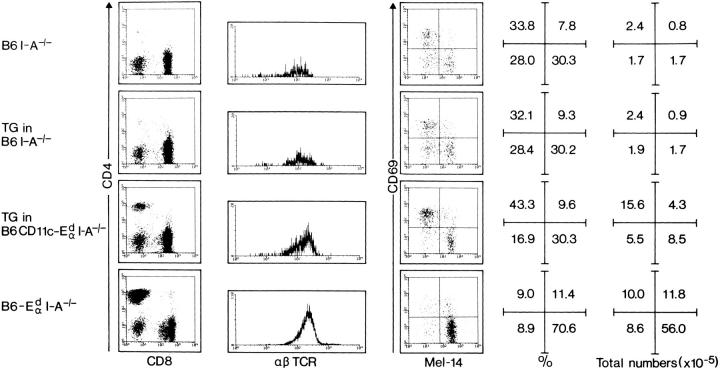
Thymic T cell development is controlled by T cell receptor (TCR)-major histocompatibility complex (MHC) interactions, whereas a further dependence of peripheral mature T cells on TCR-MHC contact has not been described so far. To study this question, CD4 T cell survival was surveyed in mice lacking MHC class II expression and in mice expressing MHC class II exclusively on dendritic cells. Since neither of these mice positively select CD4 T cells in the thymus, they were grafted with MHC class II-positive embryonic thymic tissue, which had been depleted of bone marrow derived cells. Although the thymus grafts in both hosts were repopulated with host origin thymocytes of identical phenotype and numbers, an accumulation of CD4+ T cells in peripheral lymphoid organs could only be observed in mice expressing MHC class II on dendritic cells, but not in mice that were completely MHC class II deficient. As assessed by histology, the accumulating peripheral CD4 T cells were found to be in close contact with MHC class II+ dendritic cells, suggesting that CD4 T cells need peripheral MHC class II expression for survival and that class II+ dendritic cells might play an important role for the longevity of CD4 T cells.
Figures

Similar articles
-
Murine CD4 T cells selected in a highly disparate xenogeneic porcine thymus graft do not show rapid decay in the absence of selecting MHC in the periphery.J Immunol. 2002 Dec 15;169(12):6697-710. doi: 10.4049/jimmunol.169.12.6697. J Immunol. 2002. PMID: 12471101
-
In vivo treatment of class II MHC-deficient mice with anti-TCR antibody restores the generation of circulating CD4 T cells and optimal architecture of thymic medulla.J Immunol. 2003 Oct 1;171(7):3394-400. doi: 10.4049/jimmunol.171.7.3394. J Immunol. 2003. PMID: 14500633
-
CD4+ T cells mature in the absence of MHC class I and class II expression in Ly-6A.2 transgenic mice.J Immunol. 1998 Jul 1;161(1):175-82. J Immunol. 1998. PMID: 9647222
-
Dendritic cells as stimulator cells of MHC class I-restricted immune responses.Adv Exp Med Biol. 1995;378:341-5. doi: 10.1007/978-1-4615-1971-3_76. Adv Exp Med Biol. 1995. PMID: 8526088 Review.
-
MHC-dependent survival of naïve T cells? A complicated answer to a simple question.Microbes Infect. 2002 Apr;4(5):547-54. doi: 10.1016/s1286-4579(02)01571-x. Microbes Infect. 2002. PMID: 11959510 Review.
Cited by
-
Homeostatic T cell proliferation: how far can T cells be activated to self-ligands?J Exp Med. 2000 Aug 21;192(4):F9-F14. doi: 10.1084/jem.192.4.f9. J Exp Med. 2000. PMID: 10952731 Free PMC article. Review. No abstract available.
-
Treatment of an autoimmune disease with "classical" T cell veto: a proposal.J Clin Immunol. 1999 Jul;19(4):195-202. doi: 10.1023/a:1020511928974. J Clin Immunol. 1999. PMID: 10471973 Review.
-
T cell homeostasis requires G protein-coupled receptor-mediated access to trophic signals that promote growth and inhibit chemotaxis.Eur J Immunol. 2005 Mar;35(3):786-95. doi: 10.1002/eji.200425729. Eur J Immunol. 2005. PMID: 15719365 Free PMC article.
-
Murine neonatal recent thymic emigrants are phenotypically and functionally distinct from adult recent thymic emigrants.Blood. 2009 May 28;113(22):5635-43. doi: 10.1182/blood-2008-08-173658. Epub 2009 Jan 23. Blood. 2009. PMID: 19168791 Free PMC article.
-
Partial loss of Sorting Nexin 27 resembles age- and Down syndrome-associated T cell dysfunctions.Immun Ageing. 2024 Jan 2;21(1):2. doi: 10.1186/s12979-023-00402-3. Immun Ageing. 2024. PMID: 38166948 Free PMC article.
References
-
- Jameson SC, Hogquist KA, Bevan MJ. Positive selection of thymocytes. Annu Rev Immunol. 1995;13:93–126. - PubMed
-
- Nossal GJ. Negative selection of lymphocytes. Cell. 1994;76:229–239. - PubMed
-
- Anderson G, Moore NC, Owen JJT, Jenkinson EJ. Cellular interactions in thymocyte development. Annu Rev Immunol. 1996;14:73–99. - PubMed
-
- Kontgen F, Suss G, Stewart C, Steinmetz M, Bluethmann H. Targeted disruption of the MHC class II Aa gene in C57BL/6 mice. Int Immunol. 1993;5:957–964. - PubMed
-
- Gosgrove D, Gray D, Dierich A, Kaufman J, Lemeur M, Benoist C, Mathis D. Mice lacking MHC class II molecules. Cell. 1991;66:1051–1066. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
