

Comparison of the ion channel characteristics of proapoptotic BAX and antiapoptotic BCL-2
- PMID: 9326614
- PMCID: PMC23466
- DOI: 10.1073/pnas.94.21.11357
Comparison of the ion channel characteristics of proapoptotic BAX and antiapoptotic BCL-2
Abstract
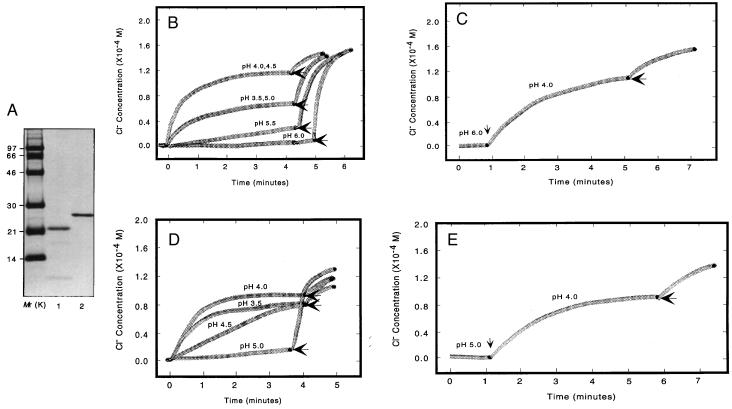
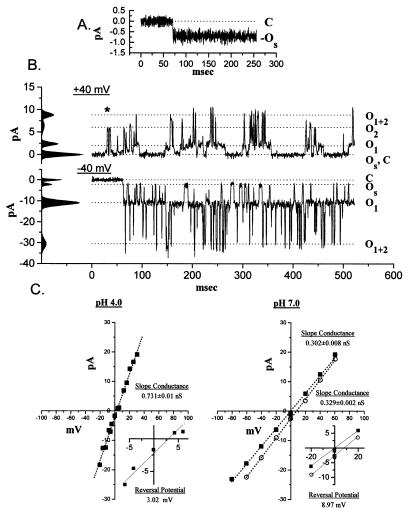
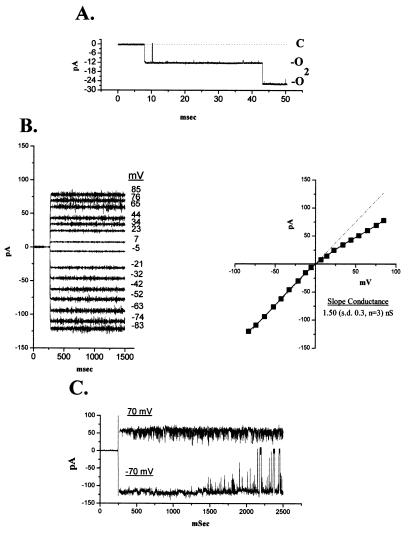
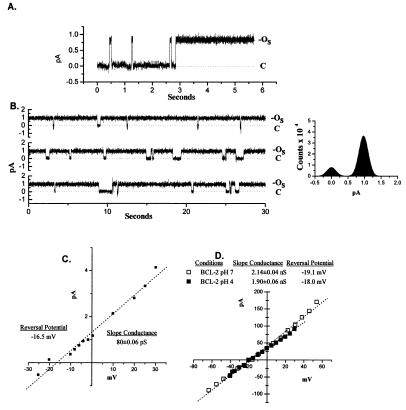
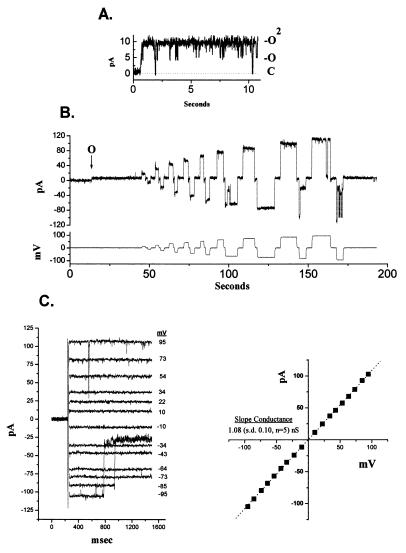
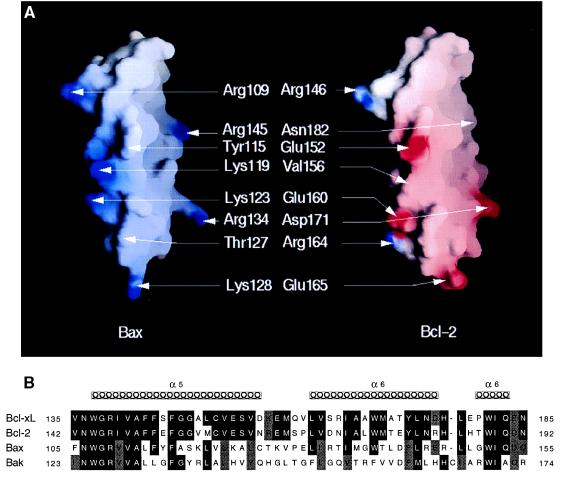
The BCL-2 family of proteins is composed of both pro- and antiapoptotic regulators, although its most critical biochemical functions remain uncertain. The structural similarity between the BCL-XL monomer and several ion-pore-forming bacterial toxins has prompted electrophysiologic studies. Both BAX and BCL-2 insert into KCl-loaded vesicles in a pH-dependent fashion and demonstrate macroscopic ion efflux. Release is maximum at approximately pH 4.0 for both proteins; however, BAX demonstrates a broader pH range of activity. Both purified proteins also insert into planar lipid bilayers at pH 4.0. Single-channel recordings revealed a minimal channel conductance for BAX of 22 pS that evolved to channel currents with at least three subconductance levels. The final, apparently stable BAX channel had a conductance of 0.731 nS at pH 4. 0 that changed to 0.329 nS when shifted to pH 7.0 but remained mildly Cl- selective and predominantly open. When BAX-incorporated lipid vesicles were fused to planar lipid bilayers at pH 7.0, a Cl--selective (PK/PCl = 0.3) 1.5-nS channel displaying mild inward rectification was noted. In contrast, BCL-2 formed mildly K+-selective (PK/PCl = 3.9) channels with a most prominent initial conductance of 80 pS that increased to 1.90 nS. Fusion of BCL-2-incorporated lipid vesicles into planar bilayers at pH 7.0 also revealed mild K+ selectivity (PK/PCl = 2.4) with a maximum conductance of 1.08 nS. BAX and BCL-2 each form channels in artificial membranes that have distinct characteristics including ion selectivity, conductance, voltage dependence, and rectification. Thus, one role of these molecules may include pore activity at selected membrane sites.
Figures
Similar articles
-
Inhibition of Bax channel-forming activity by Bcl-2.Science. 1997 Jul 18;277(5324):370-2. doi: 10.1126/science.277.5324.370. Science. 1997. PMID: 9219694
-
Channel formation by antiapoptotic protein Bcl-2.Proc Natl Acad Sci U S A. 1997 May 13;94(10):5113-8. doi: 10.1073/pnas.94.10.5113. Proc Natl Acad Sci U S A. 1997. PMID: 9144199 Free PMC article.
-
Bcl-x(L) forms an ion channel in synthetic lipid membranes.Nature. 1997 Jan 23;385(6614):353-7. doi: 10.1038/385353a0. Nature. 1997. PMID: 9002522
-
Bcl-2 family proteins as ion-channels.Cell Death Differ. 1998 May;5(5):372-80. doi: 10.1038/sj.cdd.4400365. Cell Death Differ. 1998. PMID: 10200486 Review.
-
Bax. The pro-apoptotic Bcl-2 family member, Bax.Int J Biochem Cell Biol. 1998 Jun;30(6):647-50. doi: 10.1016/s1357-2725(98)00006-5. Int J Biochem Cell Biol. 1998. PMID: 9695020 Review.
Cited by
-
Galeterone and VNPT55 induce proteasomal degradation of AR/AR-V7, induce significant apoptosis via cytochrome c release and suppress growth of castration resistant prostate cancer xenografts in vivo.Oncotarget. 2015 Sep 29;6(29):27440-60. doi: 10.18632/oncotarget.4578. Oncotarget. 2015. PMID: 26196320 Free PMC article.
-
Ligand activation of alternatively spliced fibroblast growth factor receptor-1 modulates pancreatic adenocarcinoma cell malignancy.J Gastrointest Surg. 2002 Jul-Aug;6(4):546-53. doi: 10.1016/s1091-255x(02)00036-7. J Gastrointest Surg. 2002. PMID: 12127120
-
MAC and Bcl-2 family proteins conspire in a deadly plot.Biochim Biophys Acta. 2010 Jun-Jul;1797(6-7):1231-8. doi: 10.1016/j.bbabio.2010.01.007. Epub 2010 Jan 18. Biochim Biophys Acta. 2010. PMID: 20083086 Free PMC article. Review.
-
Spatial and temporal association of Bax with mitochondrial fission sites, Drp1, and Mfn2 during apoptosis.J Cell Biol. 2002 Dec 23;159(6):931-8. doi: 10.1083/jcb.200209124. Epub 2002 Dec 23. J Cell Biol. 2002. PMID: 12499352 Free PMC article.
-
Activation of membrane-associated procaspase-3 is regulated by Bcl-2.J Cell Biol. 1999 Mar 8;144(5):915-26. doi: 10.1083/jcb.144.5.915. J Cell Biol. 1999. PMID: 10085291 Free PMC article.
References
-
- Farrow S N, Brown R. Curr Opin Gen Dev. 1996;6:45–49. - PubMed
-
- Oltvai Z N, Milliman C L, Korsmeyer S J. Cell. 1993;74:609–619. - PubMed
-
- Hockenbery D, Nunez G, Milliman C, Schreiber R D, Korsmeyer S J. Nature (London) 1990;348:334–336. - PubMed
-
- Krajewski S, Tanaka S, Takayama S, Schibler M J, Fenton W, Reed J C. Cancer Res. 1993;53:4701–4714. - PubMed
-
- Nguyen M, Millar D G, Yong V W, Korsmeyer S J, Shore G C. J Biol Chem. 1993;268:25265–25268. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
