

Phenotypic specification of hindbrain rhombomeres and the origins of rhythmic circuits in vertebrates
- PMID: 9217990
- PMCID: PMC3023276
- DOI: 10.1159/000113351
Phenotypic specification of hindbrain rhombomeres and the origins of rhythmic circuits in vertebrates
Abstract
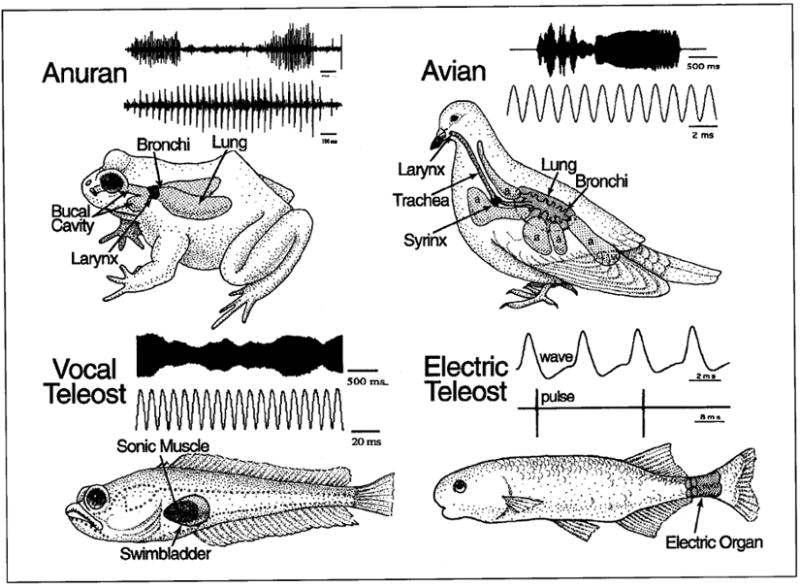
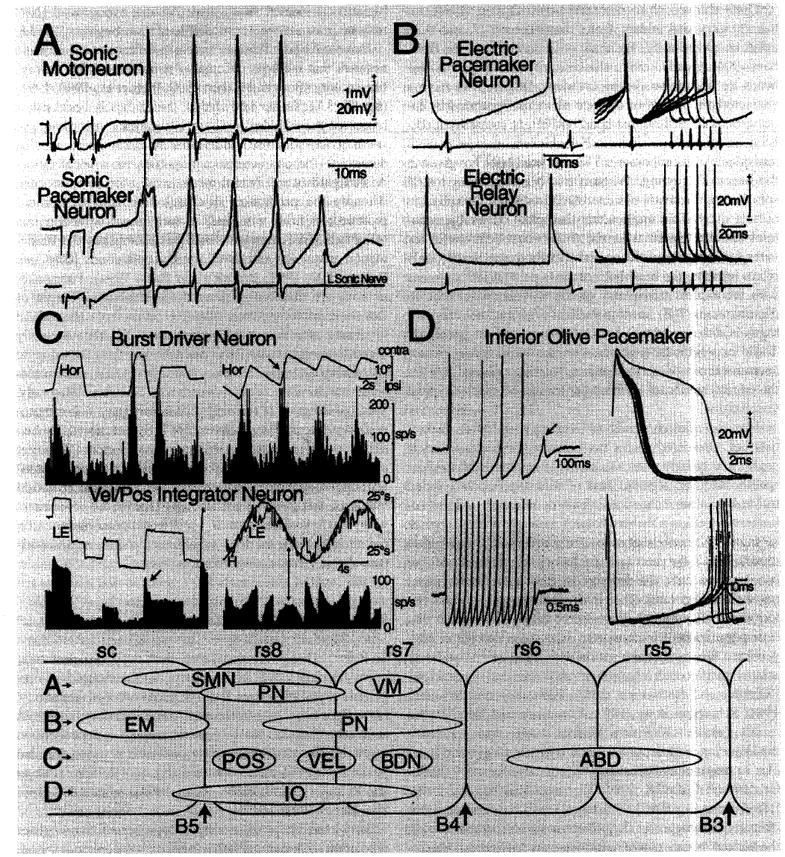
This essay considers the ontogeny and phylogeny of the cranial neural circuitry producing rhythmic behaviors in vertebrates. These behaviors are characterized by predictable temporal patterns established by a neuronal network variously referred to as either a pacemaker, neural oscillator or central pattern generator. Comparative vertebrate studies have demonstrated that the embryonic hindbrain is divided into segmented compartments called rhombomeres, each of which gives rise to a distinct complement of cranial motoneurons and, as yet, unidentified populations of interneurons. We now propose that novel rhythmic circuits were innovations associated with the adoption of cardiac and respiratory pumps during the protochordate-vertebrate transition. We further suggest that the pattern-generating circuits of more recent innovations, such as the vocal, electromotor and extraocular systems, have originated from the same Hox gene-specified compartments of the embryonic hindbrain (rhombomeres 7-8) that gave rise to rhythmically active cardiac and respiratory circuits. Lastly, we propose that the capability for pattern generation by neurons originating from rhombomeres 7 and 8 is due to their electroresponsive properties producing pacemaker oscillations, as best typified by the inferior olive which also has origins from these same hindbrain compartments and has been suggested to establish rhythmic oscillations coupled to sensorimotor function throughout the neuraxis of vertebrates.
Figures

Similar articles
-
Molecular mechanisms of pattern formation in the vertebrate hindbrain.Ciba Found Symp. 1992;165:92-102; discussion 102-7. doi: 10.1002/9780470514221.ch6. Ciba Found Symp. 1992. PMID: 1355422 Review.
-
A Hox regulatory network of hindbrain segmentation is conserved to the base of vertebrates.Nature. 2014 Oct 23;514(7523):490-3. doi: 10.1038/nature13723. Epub 2014 Sep 14. Nature. 2014. PMID: 25219855 Free PMC article.
-
A Hox gene regulatory network for hindbrain segmentation.Curr Top Dev Biol. 2020;139:169-203. doi: 10.1016/bs.ctdb.2020.03.001. Epub 2020 Apr 9. Curr Top Dev Biol. 2020. PMID: 32450960 Review.
-
Correlated patterns of neuron differentiation and Hox gene expression in the hindbrain: a comparative analysis.Brain Res Bull. 2001 Aug;55(6):683-93. doi: 10.1016/s0361-9230(01)00562-7. Brain Res Bull. 2001. PMID: 11595353 Review.
-
The vertebrate Hox gene regulatory network for hindbrain segmentation: Evolution and diversification: Coupling of a Hox gene regulatory network to hindbrain segmentation is an ancient trait originating at the base of vertebrates.Bioessays. 2016 Jun;38(6):526-38. doi: 10.1002/bies.201600010. Epub 2016 Mar 29. Bioessays. 2016. PMID: 27027928 Review.
Cited by
-
Distribution of androgen receptor mRNA expression in vocal, auditory, and neuroendocrine circuits in a teleost fish.J Comp Neurol. 2010 Feb 15;518(4):493-512. doi: 10.1002/cne.22233. J Comp Neurol. 2010. PMID: 20020540 Free PMC article.
-
Mosaic hoxb4a neuronal pleiotropism in zebrafish caudal hindbrain.PLoS One. 2009 Jun 17;4(6):e5944. doi: 10.1371/journal.pone.0005944. PLoS One. 2009. PMID: 19536294 Free PMC article.
-
Key Features of Structural and Functional Organization of Zebrafish Facial Motor Neurons Are Resilient to Disruption of Neuronal Migration.Curr Biol. 2017 Jun 19;27(12):1746-1756.e5. doi: 10.1016/j.cub.2017.05.033. Epub 2017 Jun 9. Curr Biol. 2017. PMID: 28602649 Free PMC article.
-
Non-classical 1p36 deletion in a patient with Duane retraction syndrome: case report and literature review.Mol Cytogenet. 2020 Sep 7;13:42. doi: 10.1186/s13039-020-00510-5. eCollection 2020. Mol Cytogenet. 2020. PMID: 32939224 Free PMC article.
-
Sound production in piranhas is associated with modifications of the spinal locomotor pattern.J Exp Biol. 2021 May 1;224(9):jeb242336. doi: 10.1242/jeb.242336. Epub 2021 May 4. J Exp Biol. 2021. PMID: 33942099 Free PMC article.
References
-
- Aldes LD. Subcompartmental organization of the ventral (protrusor) compartment in the hypoglossal nucleus of the rat. J Comp Neurol. 1995;353:89–108. - PubMed
-
- Baker R. A contemporary view of the phylogenetic history of eye muscles and motoneurons. In: Shimazu H, Shinoda Y, editors. Vestibular and Brain Stem Control of Eye, Head and Body Movements. Japan Scientific Societies Press; Tokyo/S. Karger, Basel: 1992. pp. 3–19.
-
- Baker R, Berthoz A. Is the prepositus hypoglossi nucleus the source of another vestibulo-ocular pathway? Brain Res. 1975;86:191–127. - PubMed
-
- Balaban E. Sex differences in sounds and their causes. In: Shorrt RV, Balaban E, editors. The Differences Between the Sexes. Cambridge University Press; Cambridge: 1994. pp. 243–273.
-
- Barbas-Henry HA, Lohman AHM. The motor nuclei and primary projections of the IXth, Xth, XIth, and XIIth cranial nerves in the monitor lizard, Varanus exanthematicus. J Comp Neurol. 1984;226:565–579. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
