

Membrane-promoted unfolding of acetylcholinesterase: a possible mechanism for insertion into the lipid bilayer
- PMID: 9096309
- PMCID: PMC20285
- DOI: 10.1073/pnas.94.7.2848
Membrane-promoted unfolding of acetylcholinesterase: a possible mechanism for insertion into the lipid bilayer
Abstract
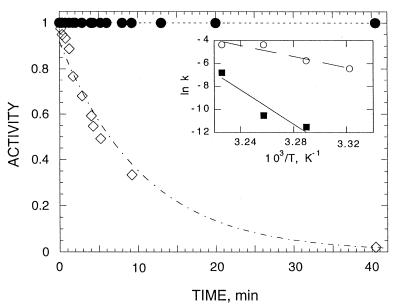
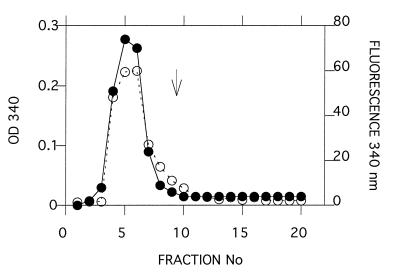
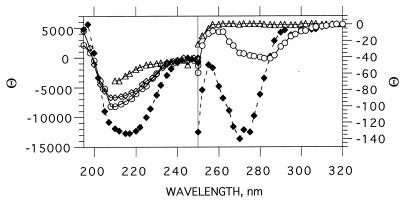
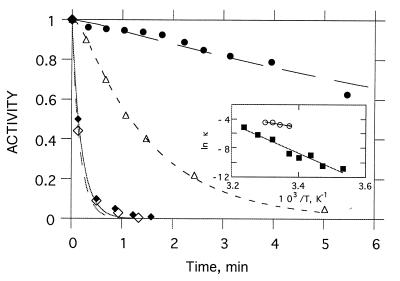
Acetylcholinesterase from Torpedo californica partially unfolds to a state with the physicochemical characteristics of a "molten globule" upon mild thermal denaturation or upon chemical modification of a single non-conserved buried cysteine residue, Cys231. The protein in this state binds tightly to liposomes. It is here shown that the rate of unfolding is greatly enhanced in the presence of unilamellar vesicles of dimyristoylphosphatidylcholine, with concomitant incorporation of the protein into the lipid bilayer. Arrhenius plots reveal that in the presence of the liposomes the energy barrier for transition from the native to the molten globule state is lowered from 145 to 47 kcal/mol. Chemical modification of Cys231 by mercuric chloride produces initially a quasinative state of Torpedo acetylcholinesterase which, at room temperature, undergoes spontaneous transition to a molten globule state with a half-life of 1-2 hr. This permitted temporal resolution of interaction of the quasi-native state with the membrane from the transition of the membrane-bound protein to the molten globule state. The data presented here suggest that either the native enzyme, or a quasi-native state with which it is in equilibrium, interacts with the liposome, which then promotes a fast transition to the membrane-bound molten globule state by lowering the energy barrier for the transition. These findings raise the possibility that the membrane itself, by lowering the energy barrier for transition to a partially unfolded state, may play an active posttranslational role in insertion and translocation of proteins in situ.
Figures
Similar articles
-
Liposome-catalyzed unfolding of acetylcholinesterase from Bungarus fasciatus.Biochemistry. 1998 Mar 31;37(13):4310-6. doi: 10.1021/bi973005q. Biochemistry. 1998. PMID: 9556345
-
Interaction of partially unfolded forms of Torpedo acetylcholinesterase with liposomes.Protein Sci. 1996 Jan;5(1):42-51. doi: 10.1002/pro.5560050106. Protein Sci. 1996. PMID: 8771195 Free PMC article.
-
Two partially unfolded states of Torpedo californica acetylcholinesterase.Protein Sci. 1996 Sep;5(9):1852-64. doi: 10.1002/pro.5560050911. Protein Sci. 1996. PMID: 8880909 Free PMC article.
-
The molten globule state of alpha-lactalbumin.FASEB J. 1996 Jan;10(1):102-9. doi: 10.1096/fasebj.10.1.8566530. FASEB J. 1996. PMID: 8566530 Review.
-
Structural energetics of the molten globule state.Proteins. 1993 Jun;16(2):115-40. doi: 10.1002/prot.340160202. Proteins. 1993. PMID: 8332604 Review.
Cited by
-
Selective and irreversible inhibitors of mosquito acetylcholinesterases for controlling malaria and other mosquito-borne diseases.PLoS One. 2009 Aug 28;4(8):e6851. doi: 10.1371/journal.pone.0006851. PLoS One. 2009. PMID: 19714254 Free PMC article.
-
Binding of lysozyme to phospholipid bilayers: evidence for protein aggregation upon membrane association.Biophys J. 2007 Jul 1;93(1):140-53. doi: 10.1529/biophysj.106.102749. Epub 2007 Apr 13. Biophys J. 2007. PMID: 17434939 Free PMC article.
-
Tertiary Plasticity Drives the Efficiency of Enterocin 7B Interactions with Lipid Membranes.J Phys Chem B. 2024 Mar 7;128(9):2100-2113. doi: 10.1021/acs.jpcb.3c08199. Epub 2024 Feb 27. J Phys Chem B. 2024. PMID: 38412510 Free PMC article.
-
Origin of the conformational heterogeneity of cardiolipin-bound cytochrome C.J Am Chem Soc. 2012 Nov 14;134(45):18713-23. doi: 10.1021/ja307426k. Epub 2012 Nov 2. J Am Chem Soc. 2012. PMID: 23066867 Free PMC article.
-
Stabilization of a metastable state of Torpedo californica acetylcholinesterase by chemical chaperones.Protein Sci. 2003 Oct;12(10):2337-47. doi: 10.1110/ps.03110703. Protein Sci. 2003. PMID: 14500892 Free PMC article.
References
-
- Isenman L, Liebow C, Rothman S. Biochim Biophys Acta. 1995;241:341–370. - PubMed
-
- Schatz G, Dobberstein B. Science. 1996;271:1519–1526. - PubMed
-
- Neupert W, Schatz G. Trends Biochem Sci. 1981;6:1–4. - PubMed
-
- Pelham H R B. Cell. 1986;46:959–961. - PubMed
-
- Verner K, Schatz G. Science. 1988;241:1307–1313. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
