

Visual motion-detection circuits in flies: small-field retinotopic elements responding to motion are evolutionarily conserved across taxa
- PMID: 8764645
- PMCID: PMC6579017
- DOI: 10.1523/JNEUROSCI.16-15-04563.1996
Visual motion-detection circuits in flies: small-field retinotopic elements responding to motion are evolutionarily conserved across taxa
Abstract
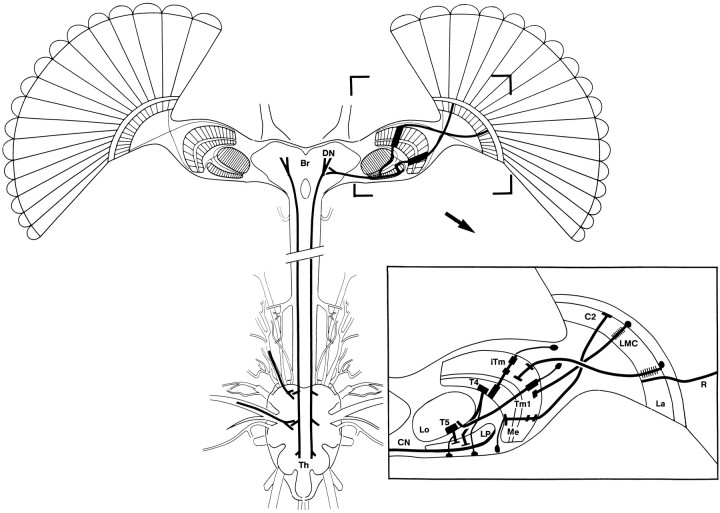
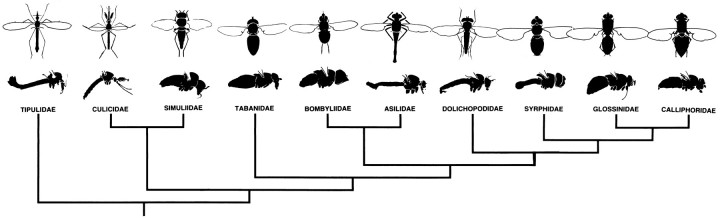
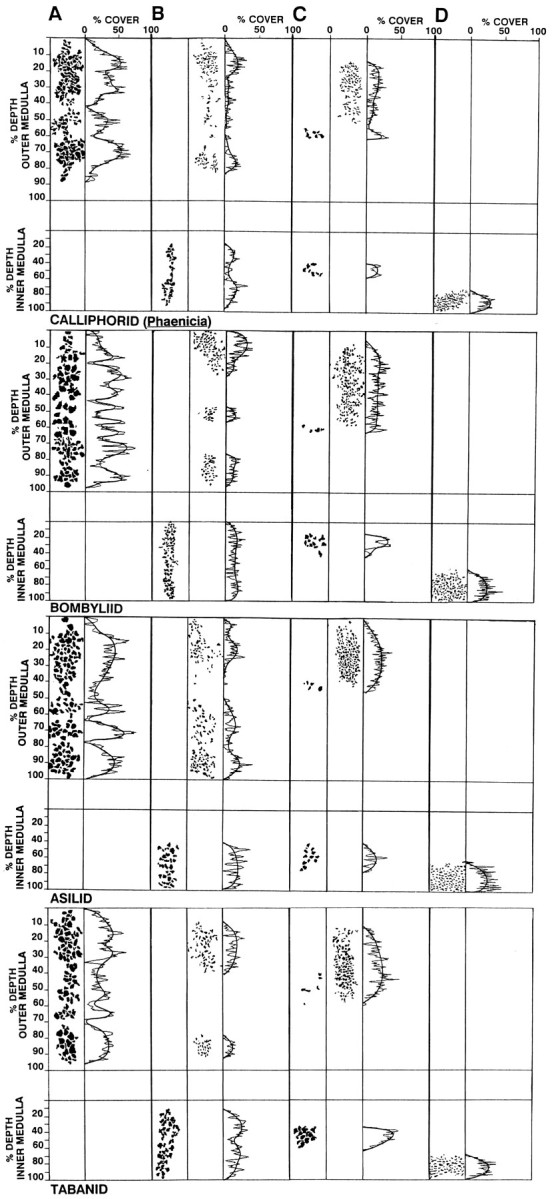
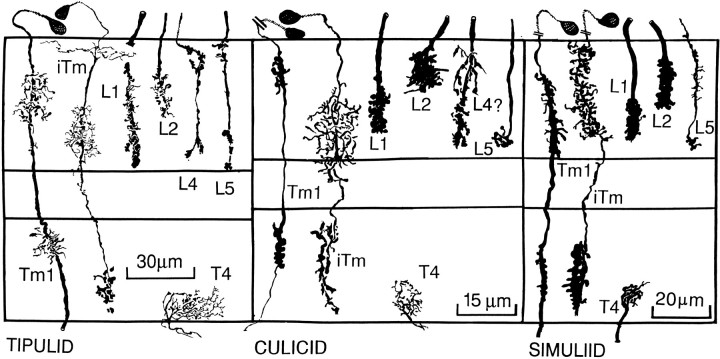
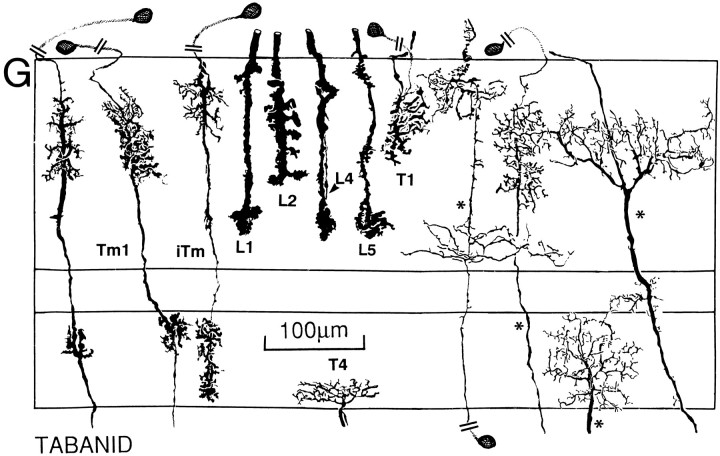
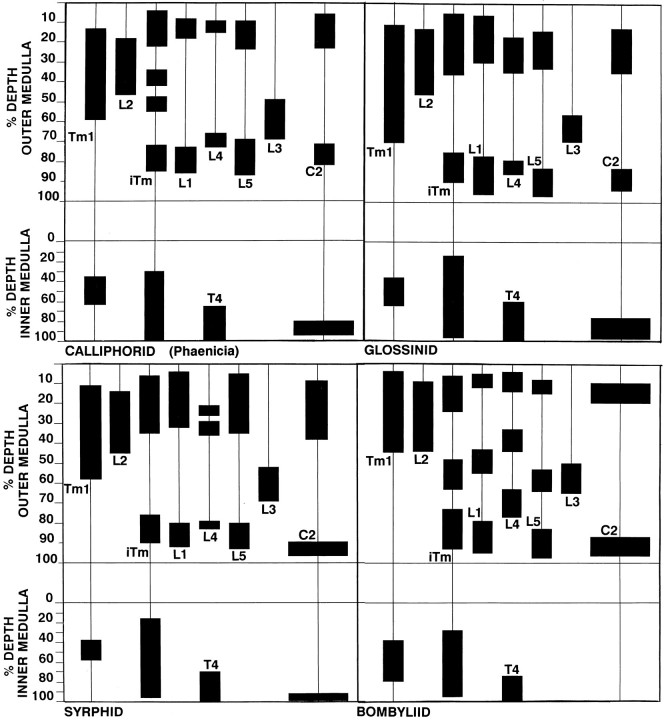
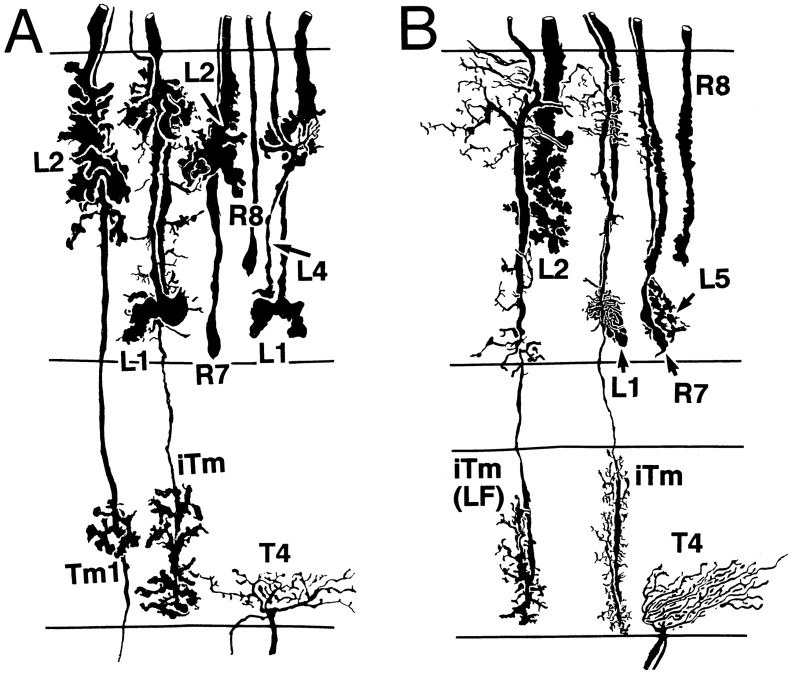
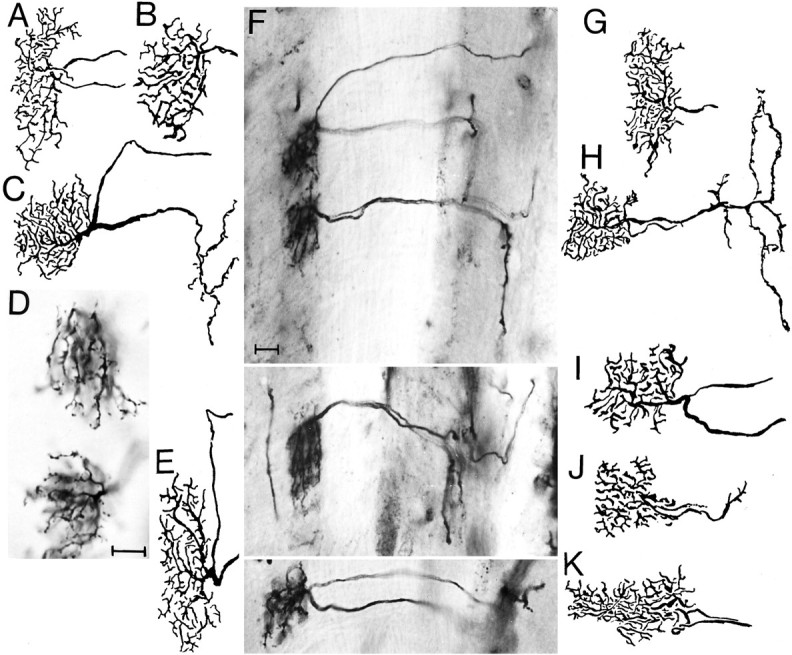
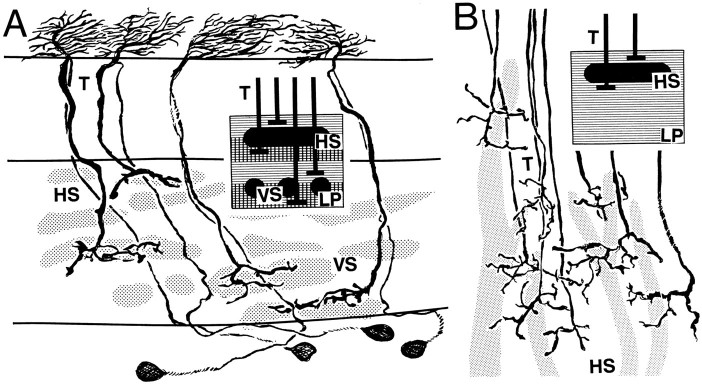
The Hassenstein-Reichardt autocorrelation model for motion computation was derived originally from studies of optomotor turning reactions of beetles and further refined from studies of houseflies. Its applicaton for explaining a variety of optokinetic behaviors in other insects assumes that neural correlates to the model are principally similar across taxa. This account examines whether this assumption is warranted. The results demonstrate that an evolutionarily conserved subset of neurons corresponds to small retinotopic neurons implicated in motion-detecting circuits that link the retina to motion-sensitive neuropils of the lobula plate. The occurrence of these neurons in basal groups suggests that they must have evolved at least 240 million years before the present time. Functional contiguity among the neurons is suggested by their having layer relationships that are independent of taxon-specific neurons, or the absence of orientation-specific motion-sensitive levels in the lobula plate.
Figures
Similar articles
-
Retinotopic pathways providing motion-selective information to the lobula from peripheral elementary motion-detecting circuits.J Comp Neurol. 2003 Mar 17;457(4):326-44. doi: 10.1002/cne.10575. J Comp Neurol. 2003. PMID: 12561074
-
Visual motion detection circuits in flies: peripheral motion computation by identified small-field retinotopic neurons.J Neurosci. 1995 Aug;15(8):5596-611. doi: 10.1523/JNEUROSCI.15-08-05596.1995. J Neurosci. 1995. PMID: 7643204 Free PMC article.
-
The computational basis of an identified neuronal circuit for elementary motion detection in dipterous insects.Vis Neurosci. 2004 Jul-Aug;21(4):567-86. doi: 10.1017/S0952523804214079. Vis Neurosci. 2004. PMID: 15579222
-
Anatomical organization of retinotopic motion-sensitive pathways in the optic lobes of flies.Microsc Res Tech. 2003 Oct 1;62(2):132-50. doi: 10.1002/jemt.10367. Microsc Res Tech. 2003. PMID: 12966499 Review.
-
Motion-detecting circuits in flies: coming into view.Annu Rev Neurosci. 2014;37:307-27. doi: 10.1146/annurev-neuro-071013-013931. Annu Rev Neurosci. 2014. PMID: 25032498 Review.
Cited by
-
Comparisons between the ON- and OFF-edge motion pathways in the Drosophila brain.Elife. 2019 Jan 9;8:e40025. doi: 10.7554/eLife.40025. Elife. 2019. PMID: 30624205 Free PMC article.
-
Cholinergic circuits integrate neighboring visual signals in a Drosophila motion detection pathway.Curr Biol. 2011 Dec 20;21(24):2077-84. doi: 10.1016/j.cub.2011.10.053. Epub 2011 Dec 1. Curr Biol. 2011. PMID: 22137471 Free PMC article.
-
Columnar cells necessary for motion responses of wide-field visual interneurons in Drosophila.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2012 May;198(5):389-95. doi: 10.1007/s00359-012-0716-3. Epub 2012 Mar 13. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2012. PMID: 22411431 Free PMC article.
-
Whole genome sequence of a long-legged fly Condylostylus longicornis from Hawai'i.Front Genet. 2023 Dec 11;14:1325213. doi: 10.3389/fgene.2023.1325213. eCollection 2023. Front Genet. 2023. PMID: 38146342 Free PMC article. No abstract available.
-
Spatio-temporal dynamics of impulse responses to figure motion in optic flow neurons.PLoS One. 2015 May 8;10(5):e0126265. doi: 10.1371/journal.pone.0126265. eCollection 2015. PLoS One. 2015. PMID: 25955416 Free PMC article.
References
-
- Bausenwein B, Fischbach K-F. Activity labeling patterns in the medulla of Drosophila melanogaster caused by motion stimuli. Cell Tissue Res. 1992a;270:25–35. - PubMed
-
- Bausenwein B, Fischbach K-F. Separation of functional pathways in the fly’s medulla: combination of 2-deoxyglucose studies with anatomical fine analysis. Singh RN. Nervous systems principles of design and function 1992b. 223 239 Wiley; New Delhi, India: Eastern.
-
- Bodian D. A new method for staining nerve fibers and nerve endings in mounted paraffin sections. Anat Rec. 1937;69:153–162.
-
- Boschek CB. On the fine structure of the peripheral retina and lamina ganglionaris of the fly, Musca domestica . Z Zellforsch Mikrosk Anat. 1971;118:369–409. - PubMed
-
- Braitenberg V. Patterns and projections in the visual system of the fly. I. Retina–lamina projections. Exp Brain Res. 1967;3:271–298. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources