

Noncanonical altPIDD1 protein: unveiling the true major translational output of the PIDD1 gene
- PMID: 39532532
- PMCID: PMC11557682
- DOI: 10.26508/lsa.202402910
Noncanonical altPIDD1 protein: unveiling the true major translational output of the PIDD1 gene
Abstract
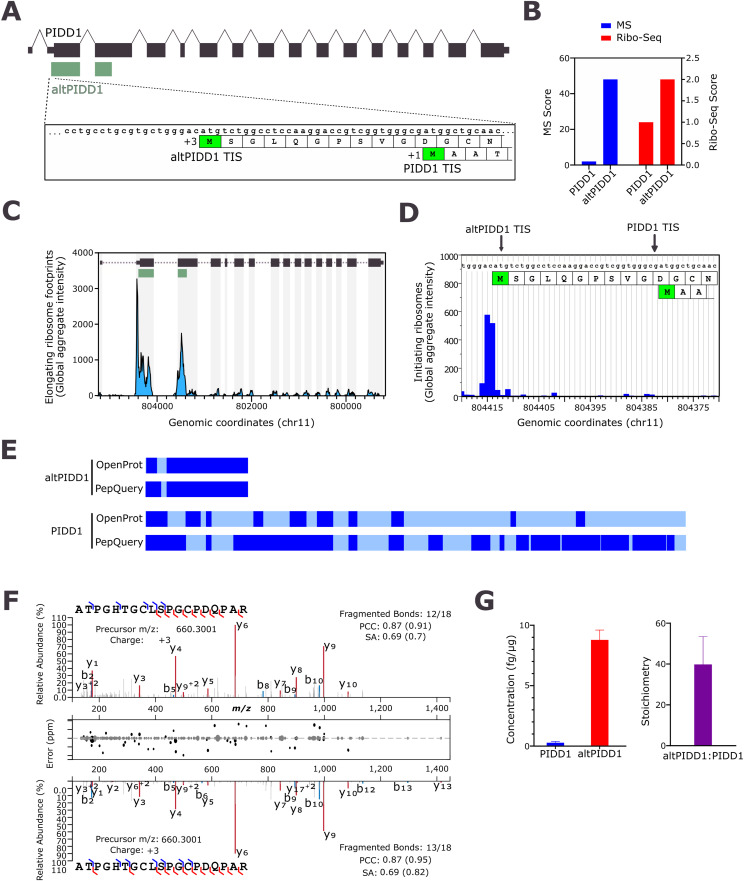
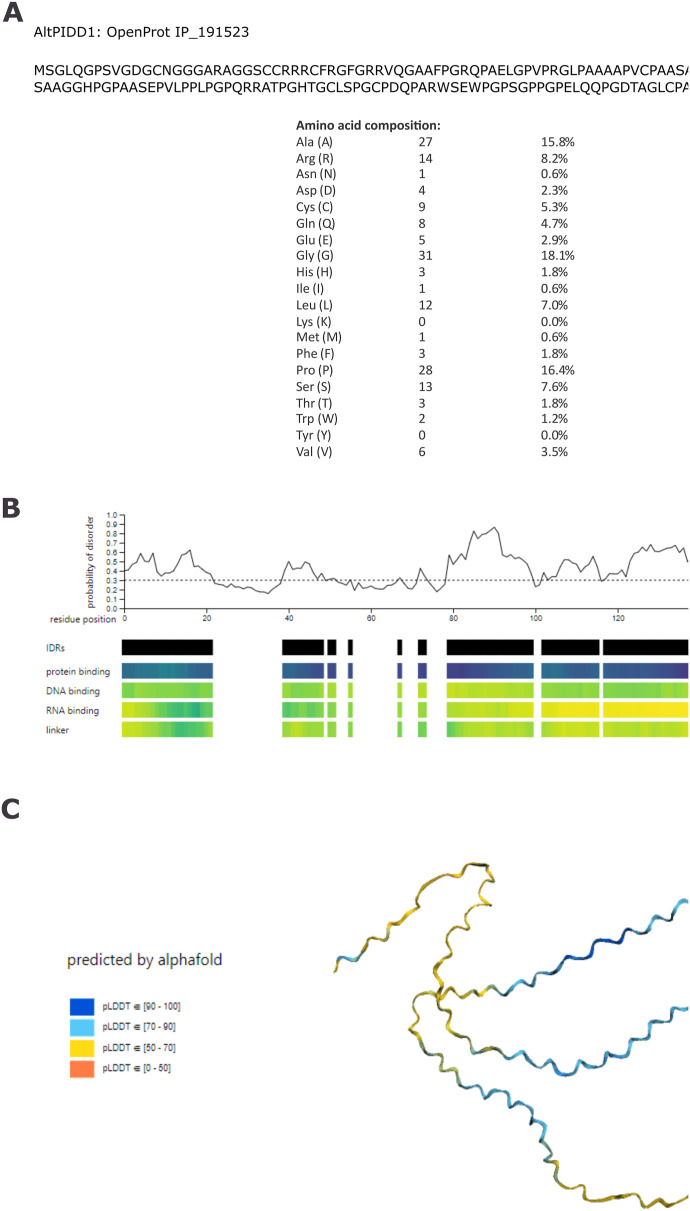
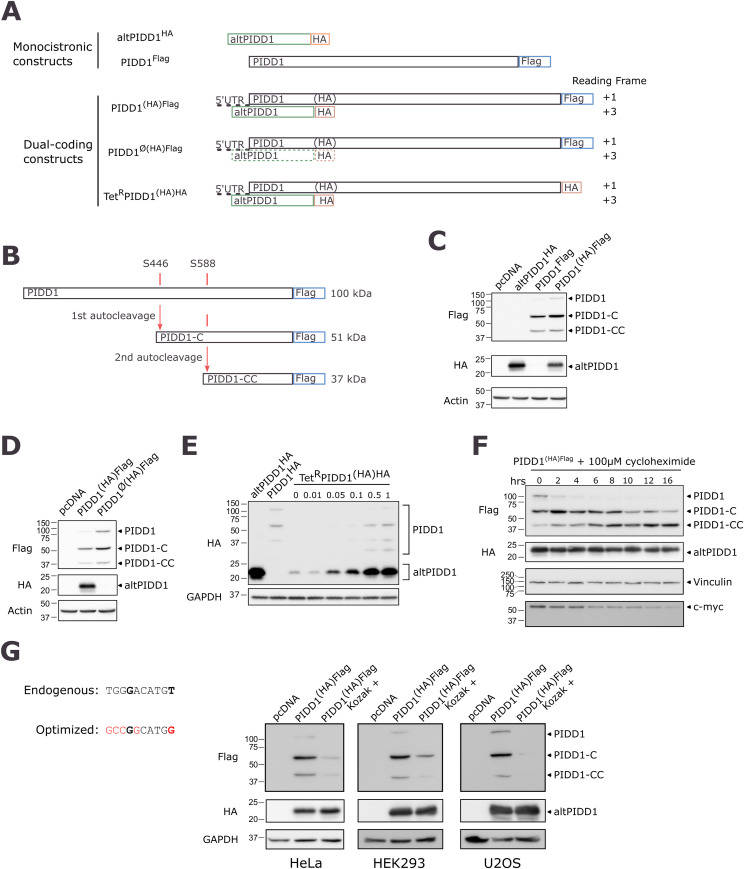
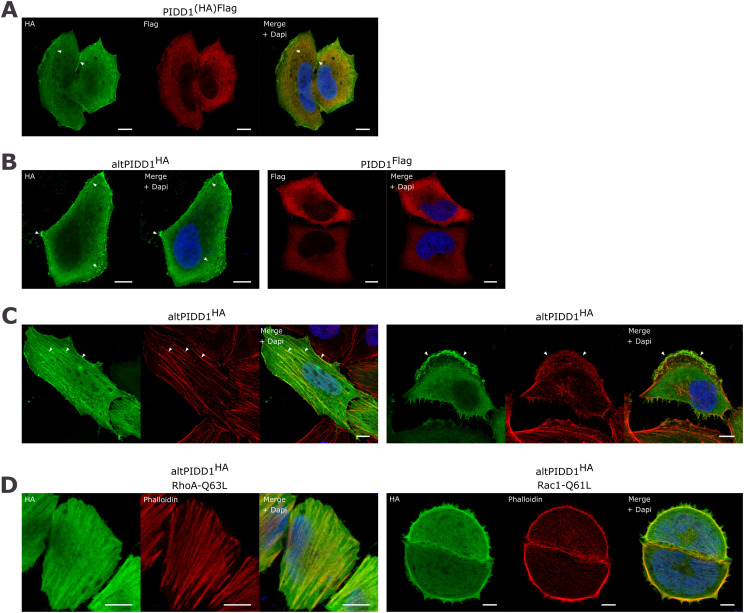
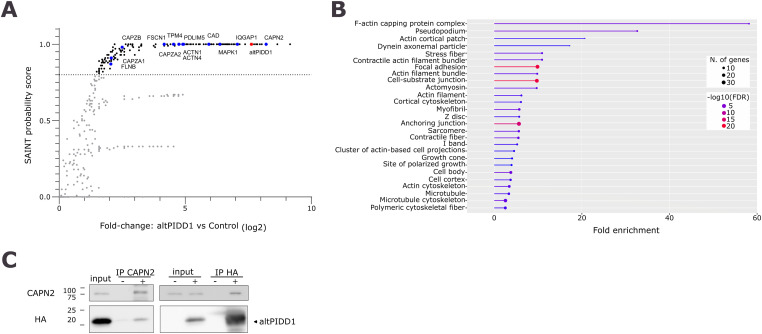
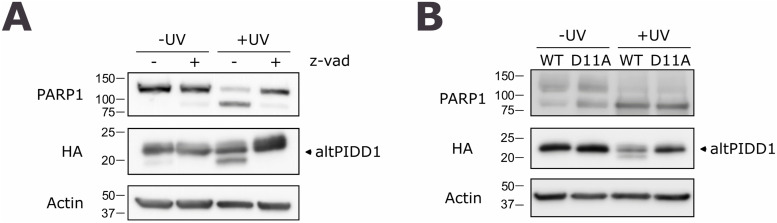
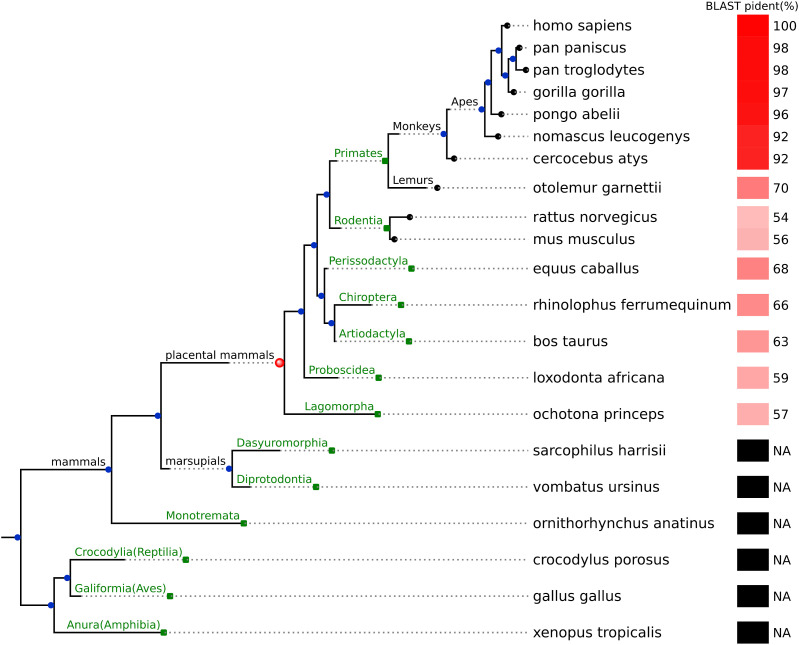
Proteogenomics has enabled the detection of novel proteins encoded in noncanonical or alternative open reading frames (altORFs) in genes already coding a reference protein. Reanalysis of proteomic and ribo-seq data revealed that the p53-induced death domain-containing protein (or PIDD1) gene encodes a second 171 amino acid protein, altPIDD1, in addition to the known 910-amino acid-long PIDD1 protein. The two ORFs overlap almost completely, and the translation initiation site of altPIDD1 is located upstream of PIDD1. AltPIDD1 has more translational and protein level evidence than PIDD1 across various cell lines and tissues. In HEK293 cells, the altPIDD1 to PIDD1 ratio is 40 to 1, as measured with isotope-labeled (heavy) peptides and targeted proteomics. AltPIDD1 localizes to cytoskeletal structures labeled with phalloidin and interacts with cytoskeletal proteins. Unlike most noncanonical proteins, altPIDD1 is not evolutionarily young but emerged in placental mammals. Overall, we identify PIDD1 as a dual-coding gene, with altPIDD1, not the annotated protein, being the primary product of translation.
© 2024 Comtois et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Impact of residual disease as a prognostic factor for survival in women with advanced epithelial ovarian cancer after primary surgery.Cochrane Database Syst Rev. 2022 Sep 26;9(9):CD015048. doi: 10.1002/14651858.CD015048.pub2. Cochrane Database Syst Rev. 2022. PMID: 36161421 Free PMC article. Review.
-
Pharmacological treatments in panic disorder in adults: a network meta-analysis.Cochrane Database Syst Rev. 2023 Nov 28;11(11):CD012729. doi: 10.1002/14651858.CD012729.pub3. Cochrane Database Syst Rev. 2023. PMID: 38014714 Free PMC article. Review.
-
Trends in Surgical and Nonsurgical Aesthetic Procedures: A 14-Year Analysis of the International Society of Aesthetic Plastic Surgery-ISAPS.Aesthetic Plast Surg. 2024 Oct;48(20):4217-4227. doi: 10.1007/s00266-024-04260-2. Epub 2024 Aug 5. Aesthetic Plast Surg. 2024. PMID: 39103642 Review.
-
Comparison of Two Modern Survival Prediction Tools, SORG-MLA and METSSS, in Patients With Symptomatic Long-bone Metastases Who Underwent Local Treatment With Surgery Followed by Radiotherapy and With Radiotherapy Alone.Clin Orthop Relat Res. 2024 Dec 1;482(12):2193-2208. doi: 10.1097/CORR.0000000000003185. Epub 2024 Jul 23. Clin Orthop Relat Res. 2024. PMID: 39051924
Cited by
-
High-quality peptide evidence for annotating non-canonical open reading frames as human proteins.bioRxiv [Preprint]. 2024 Sep 9:2024.09.09.612016. doi: 10.1101/2024.09.09.612016. bioRxiv. 2024. PMID: 39314370 Free PMC article. Preprint.
References
-
- Burigotto M, Mattivi A, Migliorati D, Magnani G, Valentini C, Roccuzzo M, Offterdinger M, Pizzato M, Schmidt A, Villunger A, et al. (2021) Centriolar distal appendages activate the centrosome-PIDDosome-p53 signalling axis via ANKRD26. EMBO J 40: e104844. 10.15252/embj.2020104844 - DOI - PMC - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous