

Genome-guided isolation of the hyperthermophilic aerobe Fervidibacter sacchari reveals conserved polysaccharide metabolism in the Armatimonadota
- PMID: 39496591
- PMCID: PMC11535203
- DOI: 10.1038/s41467-024-53784-3
Genome-guided isolation of the hyperthermophilic aerobe Fervidibacter sacchari reveals conserved polysaccharide metabolism in the Armatimonadota
Abstract
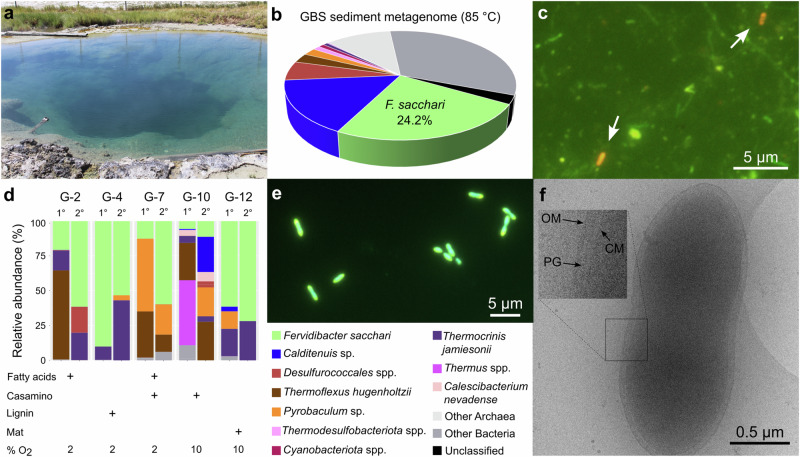
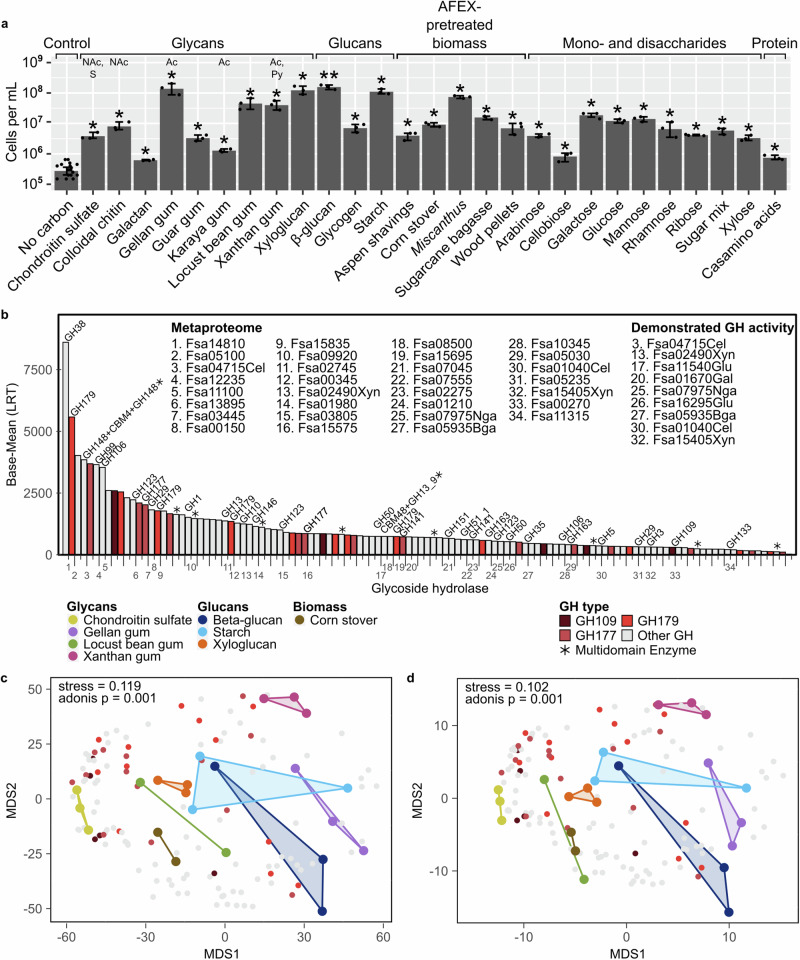
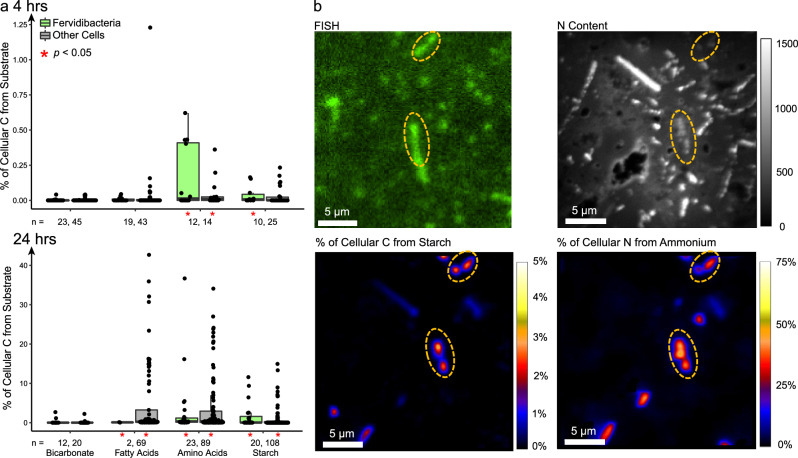
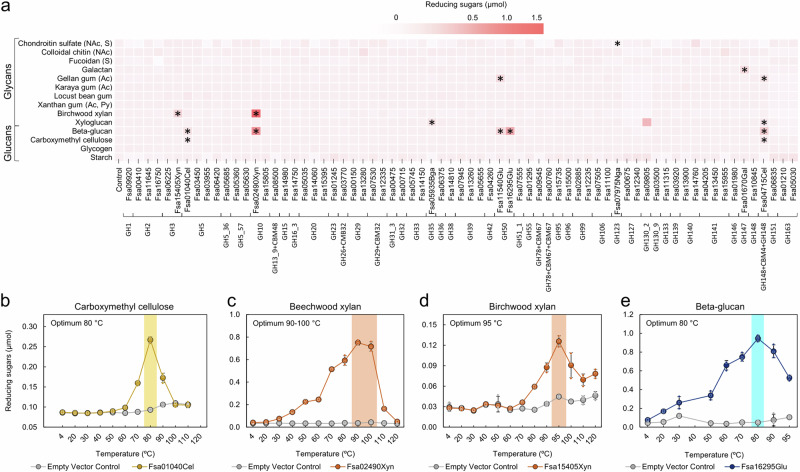
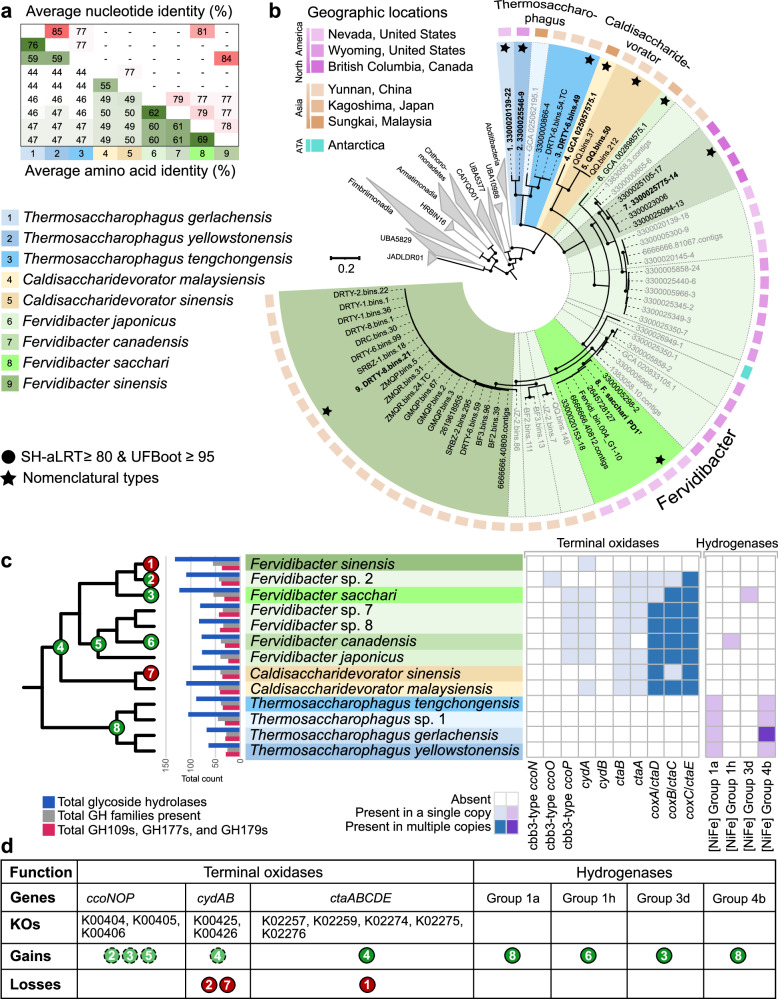
Few aerobic hyperthermophilic microorganisms degrade polysaccharides. Here, we describe the genome-enabled enrichment and optical tweezer-based isolation of an aerobic polysaccharide-degrading hyperthermophile, Fervidibacter sacchari, previously ascribed to candidate phylum Fervidibacteria. F. sacchari uses polysaccharides and monosaccharides for growth at 65-87.5 °C and expresses 191 carbohydrate-active enzymes (CAZymes) according to RNA-Seq and proteomics, including 31 with unusual glycoside hydrolase domains (GH109, GH177, GH179). Fluorescence in-situ hybridization and nanoscale secondary ion mass spectrometry confirmed rapid assimilation of 13C-starch in spring sediments. Purified GHs were optimally active at 80-100 °C on ten different polysaccharides. Finally, we propose reassigning Fervidibacteria as a class within phylum Armatimonadota, along with 18 other species, and show that a high number and diversity of CAZymes is a hallmark of the phylum, in both aerobic and anaerobic lineages. Our study establishes Fervidibacteria as hyperthermophilic polysaccharide degraders in terrestrial geothermal springs and suggests a broad role for Armatimonadota in polysaccharide catabolism.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Biochemical characterization of Fsa16295Glu from "Fervidibacter sacchari," the first hyperthermophilic GH50 with β-1,3-endoglucanase activity and founding member of the subfamily GH50_3.Front Microbiol. 2024 Apr 25;15:1355444. doi: 10.3389/fmicb.2024.1355444. eCollection 2024. Front Microbiol. 2024. PMID: 38725686 Free PMC article.
-
Comparative Genomics of Rumen Butyrivibrio spp. Uncovers a Continuum of Polysaccharide-Degrading Capabilities.Appl Environ Microbiol. 2019 Dec 13;86(1):e01993-19. doi: 10.1128/AEM.01993-19. Print 2019 Dec 13. Appl Environ Microbiol. 2019. PMID: 31653790 Free PMC article.
-
Genomic potential for polysaccharide deconstruction in bacteria.Appl Environ Microbiol. 2015 Feb;81(4):1513-19. doi: 10.1128/AEM.03718-14. Appl Environ Microbiol. 2015. PMID: 25527556 Free PMC article.
-
Interrogating gut bacterial genomes for discovery of novel carbohydrate degrading enzymes.Curr Opin Chem Biol. 2018 Dec;47:126-133. doi: 10.1016/j.cbpa.2018.09.012. Epub 2018 Oct 14. Curr Opin Chem Biol. 2018. PMID: 30326425 Review.
-
Effect of multimodularity and spatial organization of glycoside hydrolases on catalysis.Essays Biochem. 2023 Apr 18;67(3):629-638. doi: 10.1042/EBC20220167. Essays Biochem. 2023. PMID: 36866571 Review.
References
-
- Seidi, F. et al. Crystalline polysaccharides: a review. Carbohydr. Polym.275, 118624 (2022). - PubMed
-
- Blumer-Schuette, S. E., Kataeva, I., Westpheling, J., Adams, M. W. & Kelly, R. M. Extremely thermophilic microorganisms for biomass conversion: status and prospects. Curr. Opin. Biotechnol.19, 210–217 (2008). - PubMed
-
- Brochier-Armanet, C., Talla, E. & Gribaldo, S. The multiple evolutionary histories of dioxygen reductases: implications for the origin and evolution of aerobic respiration. Mol. Biol. Evol.26, 285–297 (2009). - PubMed
-
- Murali, R., Hemp, J. & Gennis, R. B. Evolution of quinol oxidation within the heme‑copper oxidoreductase superfamily. Biochim. Biophys. Acta (BBA) - Bioenerg.1863, 148907 (2022). - PubMed
MeSH terms
Substances
Grants and funding
- 91951205/National Natural Science Foundation of China (National Science Foundation of China)
- 80NNSC17KO548/National Aeronautics and Space Administration (NASA)
- 80NSSC20M0043/Nevada Space Grant Consortium (Nevada NASA Space Grant Consortium)
- P20 GM103440/GM/NIGMS NIH HHS/United States
- GM103440/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
