

A noncanonical GTPase signaling mechanism controls exit from mitosis in budding yeast
- PMID: 39475649
- PMCID: PMC11551315
- DOI: 10.1073/pnas.2413873121
A noncanonical GTPase signaling mechanism controls exit from mitosis in budding yeast
Abstract
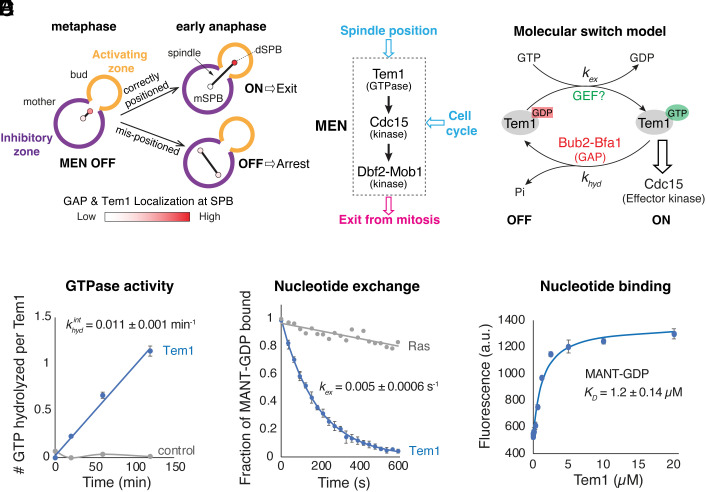
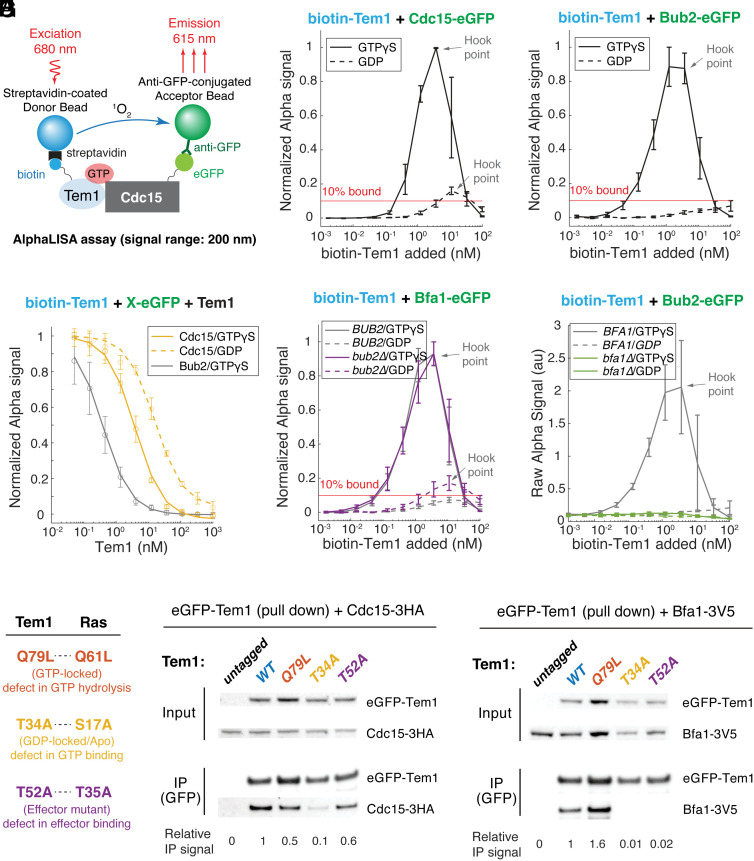
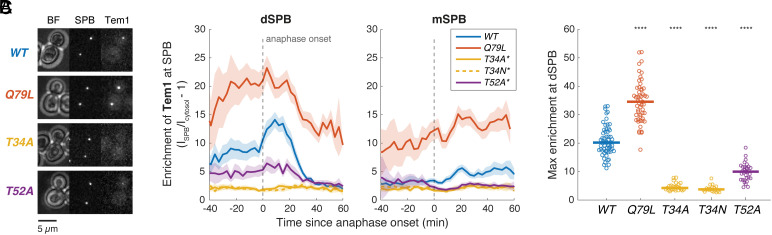
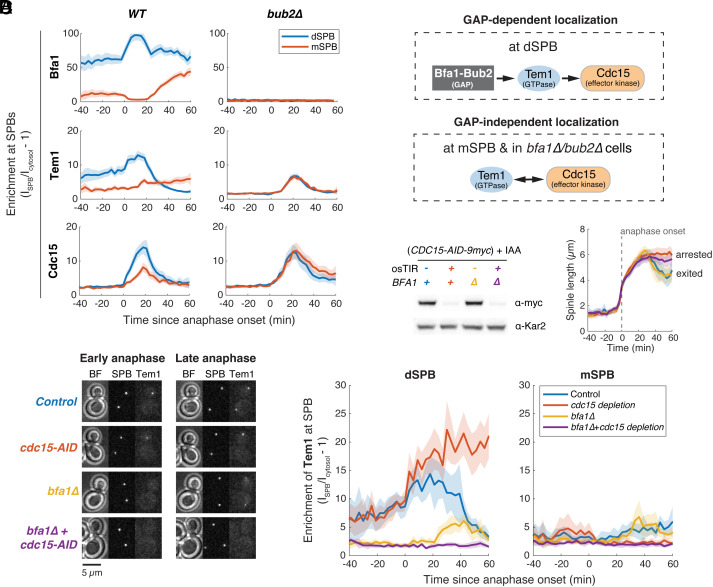
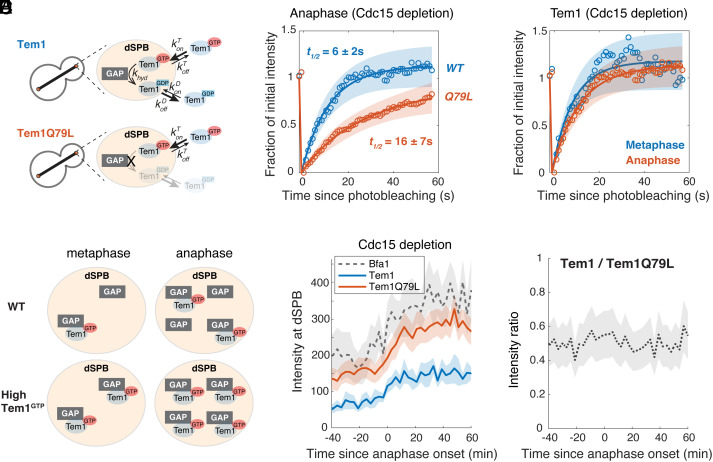
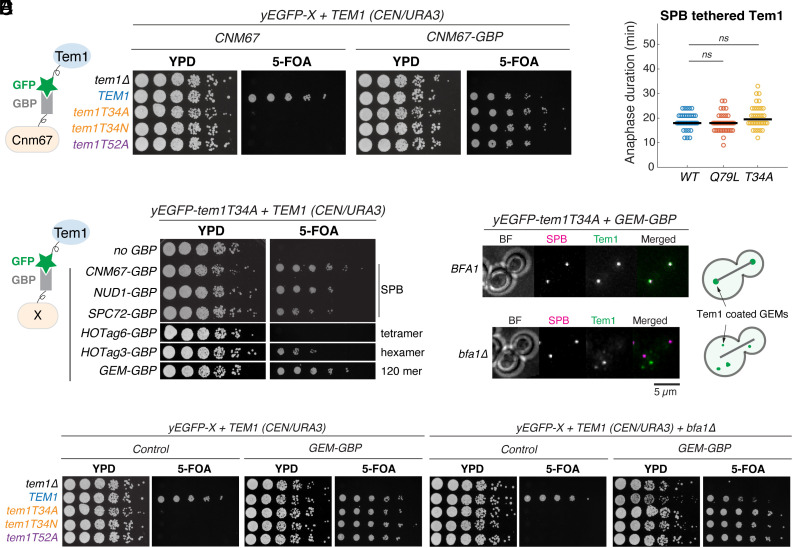
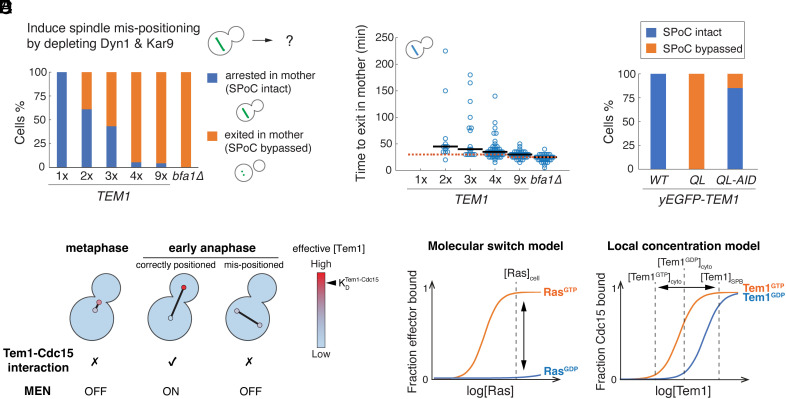
In the budding yeast Saccharomyces cerevisiae, exit from mitosis is coupled to spindle position to ensure successful genome partitioning between mother and daughter cells. This coupling occurs through a GTPase signaling cascade known as the mitotic exit network (MEN). The MEN senses spindle position via a Ras-like GTPase Tem1 which localizes to the spindle pole bodies (SPBs, yeast equivalent of centrosomes) during anaphase and signals to its effector protein kinase Cdc15. How Tem1 couples the status of spindle position to MEN activation is not fully understood. Here, we show that Cdc15 has a relatively weak preference for Tem1GTP and Tem1's nucleotide state does not change upon MEN activation. Instead, we find that Tem1's nucleotide cycle establishes a localization-based concentration difference in the cell where only Tem1GTP is recruited to the SPB, and spindle position regulates the MEN by controlling Tem1 localization to the SPB. SPB localization of Tem1 primarily functions to promote Tem1-Cdc15 interaction for MEN activation by increasing the effective concentration of Tem1. Consistent with this model, we demonstrate that artificially tethering Tem1 to the SPB or concentrating Tem1 in the cytoplasm with genetically encoded multimeric nanoparticles could bypass the requirement of Tem1GTP and correct spindle position for MEN activation. This localization/concentration-based GTPase signaling mechanism for Tem1 differs from the canonical Ras-like GTPase signaling paradigm and is likely relevant to other localization-based signaling scenarios.
Keywords: GTPase signaling; cell cycle control; mitotic exit network; spindle position checkpoint.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
Update of
-
A noncanonical GTPase signaling mechanism controls exit from mitosis in budding yeast.bioRxiv [Preprint]. 2024 Jul 4:2024.05.16.594582. doi: 10.1101/2024.05.16.594582. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 Nov 5;121(45):e2413873121. doi: 10.1073/pnas.2413873121 PMID: 38798491 Free PMC article. Updated. Preprint.
Similar articles
-
A noncanonical GTPase signaling mechanism controls exit from mitosis in budding yeast.bioRxiv [Preprint]. 2024 Jul 4:2024.05.16.594582. doi: 10.1101/2024.05.16.594582. bioRxiv. 2024. Update in: Proc Natl Acad Sci U S A. 2024 Nov 5;121(45):e2413873121. doi: 10.1073/pnas.2413873121 PMID: 38798491 Free PMC article. Updated. Preprint.
-
Spindle pole bodies function as signal amplifiers in the Mitotic Exit Network.Mol Biol Cell. 2020 Apr 15;31(9):906-916. doi: 10.1091/mbc.E19-10-0584. Epub 2020 Feb 19. Mol Biol Cell. 2020. PMID: 32074005 Free PMC article.
-
Cdc15 integrates Tem1 GTPase-mediated spatial signals with Polo kinase-mediated temporal cues to activate mitotic exit.Genes Dev. 2011 Sep 15;25(18):1943-54. doi: 10.1101/gad.17257711. Genes Dev. 2011. PMID: 21937712 Free PMC article.
-
Coupling spindle position with mitotic exit in budding yeast: The multifaceted role of the small GTPase Tem1.Small GTPases. 2015 Oct 2;6(4):196-201. doi: 10.1080/21541248.2015.1109023. Small GTPases. 2015. PMID: 26507466 Free PMC article. Review.
-
Mitotic exit control: a space and time odyssey.Curr Biol. 2011 Oct 25;21(20):R857-9. doi: 10.1016/j.cub.2011.09.023. Curr Biol. 2011. PMID: 22032192 Review.
References
-
- Wennerberg K., The Ras superfamily at a glance. J. Cell Sci. 118, 843–846 (2005). - PubMed
-
- Vetter I. R., Wittinghofer A., The guanine nucleotide-binding switch in three dimensions. Science 294, 1299–1304 (2001). - PubMed
-
- Bos J. L., Rehmann H., Ittinghofer A., GEFs and GAPs: Critical elements in the control of small G proteins. Cell 129, 865–877 (2007). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
