

Ancestral neural circuits potentiate the origin of a female sexual behavior in Drosophila
- PMID: 39468043
- PMCID: PMC11519493
- DOI: 10.1038/s41467-024-53610-w
Ancestral neural circuits potentiate the origin of a female sexual behavior in Drosophila
Abstract
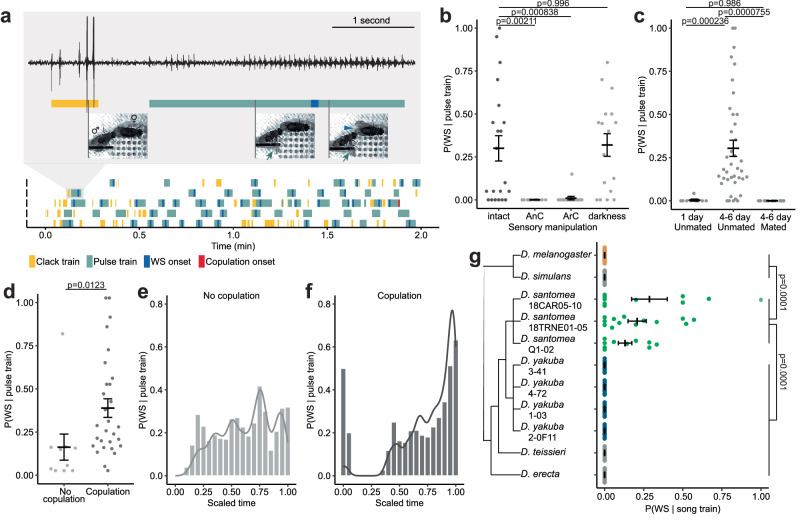
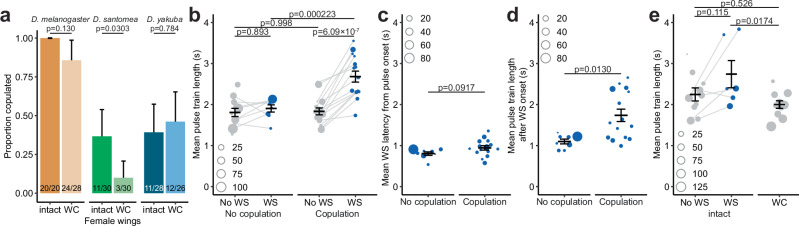
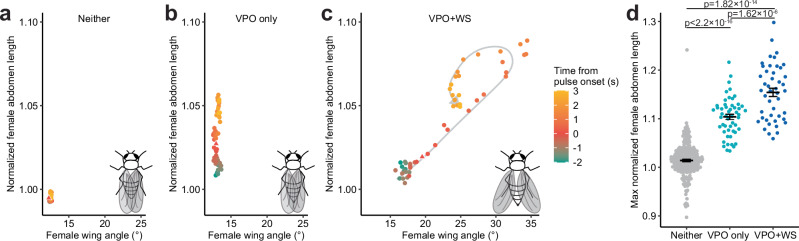
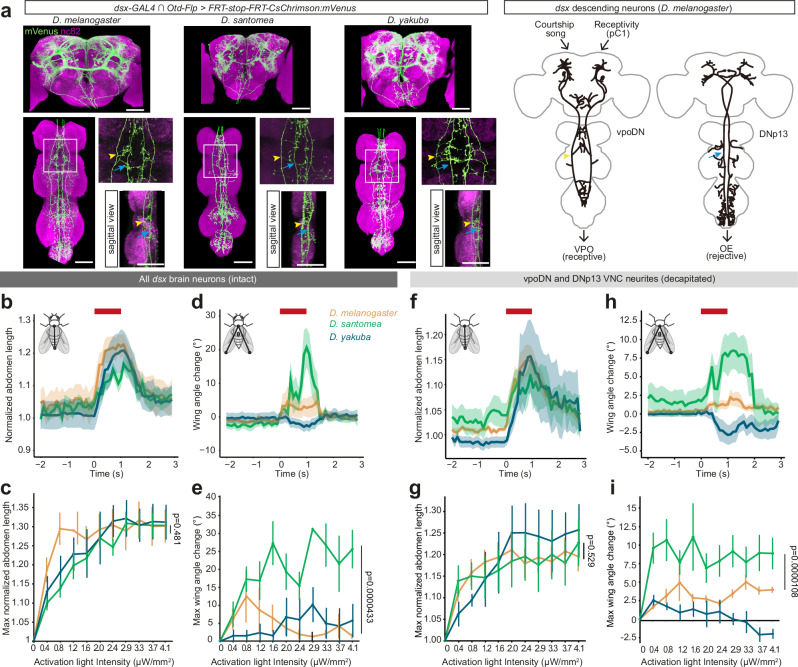
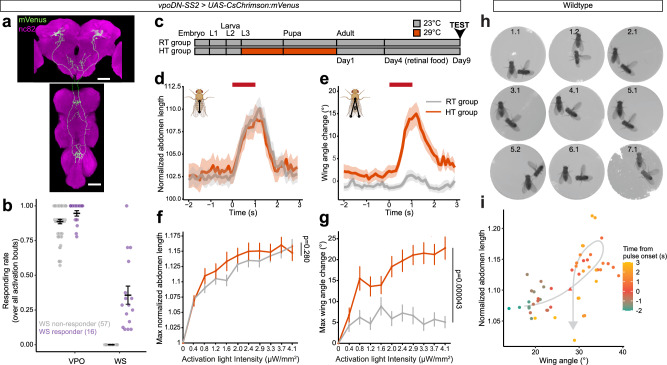
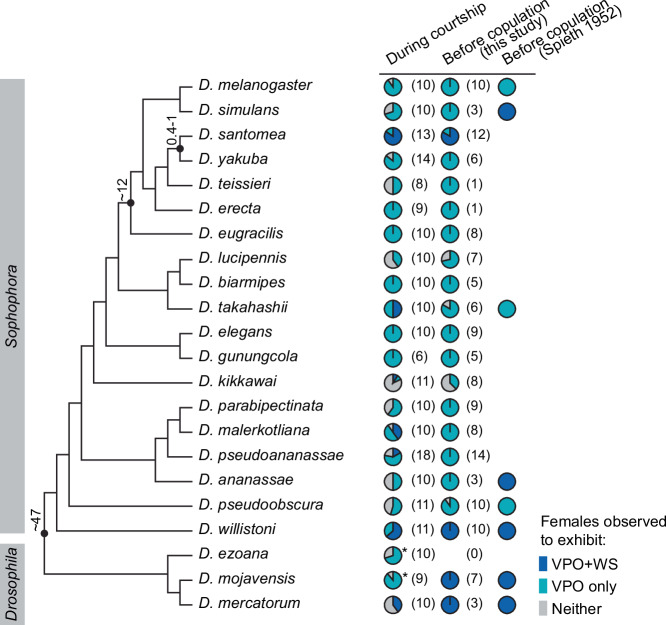
Courtship interactions are remarkably diverse in form and complexity among species. How neural circuits evolve to encode new behaviors that are functionally integrated into these dynamic social interactions is unknown. Here we report a recently originated female sexual behavior in the island endemic Drosophila species D. santomea, where females signal receptivity to male courtship songs by spreading their wings, which in turn promotes prolonged songs in courting males. Copulation success depends on this female signal and correlates with males' ability to adjust his singing in such a social feedback loop. Functional comparison of sexual circuitry across species suggests that a pair of descending neurons, which integrates male song stimuli and female internal state to control a conserved female abdominal behavior, drives wing spreading in D. santomea. This co-option occurred through the refinement of a pre-existing, plastic circuit that can be optogenetically activated in an outgroup species. Combined, our results show that the ancestral potential of a socially-tuned key circuit node to engage the wing motor circuit facilitates the expression of a new female behavior in appropriate sensory and motivational contexts. More broadly, our work provides insights into the evolution of social behaviors, particularly female behaviors, and the underlying neural mechanisms.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Ancestral neural circuits potentiate the origin of a female sexual behavior.bioRxiv [Preprint]. 2023 Dec 7:2023.12.05.570174. doi: 10.1101/2023.12.05.570174. bioRxiv. 2023. Update in: Nat Commun. 2024 Oct 28;15(1):9210. doi: 10.1038/s41467-024-53610-w. PMID: 38106147 Free PMC article. Updated. Preprint.
Similar articles
-
Ancestral neural circuits potentiate the origin of a female sexual behavior.bioRxiv [Preprint]. 2023 Dec 7:2023.12.05.570174. doi: 10.1101/2023.12.05.570174. bioRxiv. 2023. Update in: Nat Commun. 2024 Oct 28;15(1):9210. doi: 10.1038/s41467-024-53610-w. PMID: 38106147 Free PMC article. Updated. Preprint.
-
Nested neural circuits generate distinct acoustic signals during Drosophila courtship.Curr Biol. 2024 Feb 26;34(4):808-824.e6. doi: 10.1016/j.cub.2024.01.015. Epub 2024 Jan 30. Curr Biol. 2024. PMID: 38295797
-
Neural Evolution of Context-Dependent Fly Song.Curr Biol. 2019 Apr 1;29(7):1089-1099.e7. doi: 10.1016/j.cub.2019.02.019. Epub 2019 Mar 14. Curr Biol. 2019. PMID: 30880014
-
Drosophila female courtship and mating behaviors: sensory signals, genes, neural structures and evolution.Curr Opin Neurobiol. 2010 Dec;20(6):764-9. doi: 10.1016/j.conb.2010.09.007. Epub 2010 Oct 8. Curr Opin Neurobiol. 2010. PMID: 20934322 Review.
-
Neurogenetics of courtship and mating in Drosophila.Adv Genet. 2008;62:67-184. doi: 10.1016/S0065-2660(08)00603-2. Adv Genet. 2008. PMID: 19010254 Review.
Cited by
-
Temperature affects conspecific and heterospecific mating rates in Drosophila.bioRxiv [Preprint]. 2024 Oct 28:2024.10.28.620639. doi: 10.1101/2024.10.28.620639. bioRxiv. 2024. PMID: 39553966 Free PMC article. Preprint.
-
Comparative connectomics of the descending and ascending neurons of the Drosophila nervous system: stereotypy and sexual dimorphism.bioRxiv [Preprint]. 2024 Jun 28:2024.06.04.596633. doi: 10.1101/2024.06.04.596633. bioRxiv. 2024. PMID: 38895426 Free PMC article. Preprint.
References
-
- Sturtevant, A. H. Experiments on sex recognition and the problem of sexual selection in Drosoophilia. J. Exp. Psychol. Anim. Behav. Process.5, 351 (1915).
-
- Greenspan, R. J. & Ferveur, J. F. Courtship in Drosophila. Annu. Rev. Genet.34, 205–232 (2000). - PubMed
-
- Egnor, S. R. & Seagraves, K. M. The contribution of ultrasonic vocalizations to mouse courtship. Curr. Opin. Neurobiol.38, 1–5 (2016). - PubMed
-
- Perkes, A., White, D., Wild, J. M. & Schmidt, M. Female Songbirds: The unsung drivers of courtship behavior and its neural substrates. Behav. Process.163, 60–70 (2019). - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
