

CD4 downregulation precedes Env expression and protects HIV-1-infected cells from ADCC mediated by non-neutralizing antibodies
- PMID: 39373535
- PMCID: PMC11559134
- DOI: 10.1128/mbio.01827-24
CD4 downregulation precedes Env expression and protects HIV-1-infected cells from ADCC mediated by non-neutralizing antibodies
Abstract
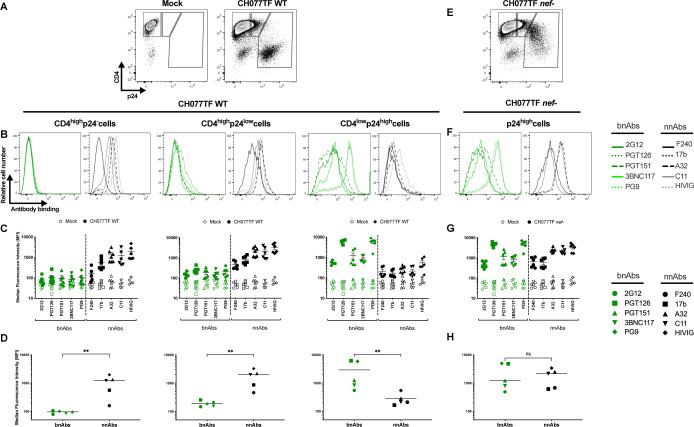
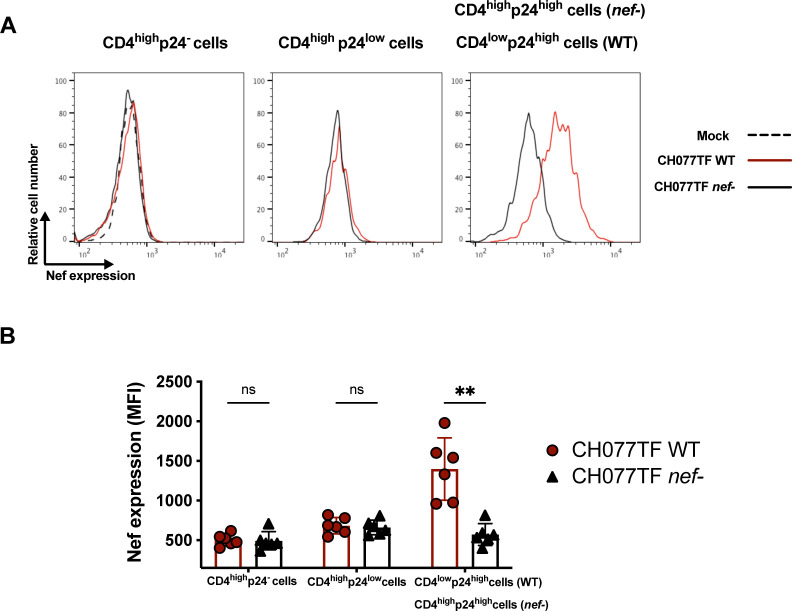
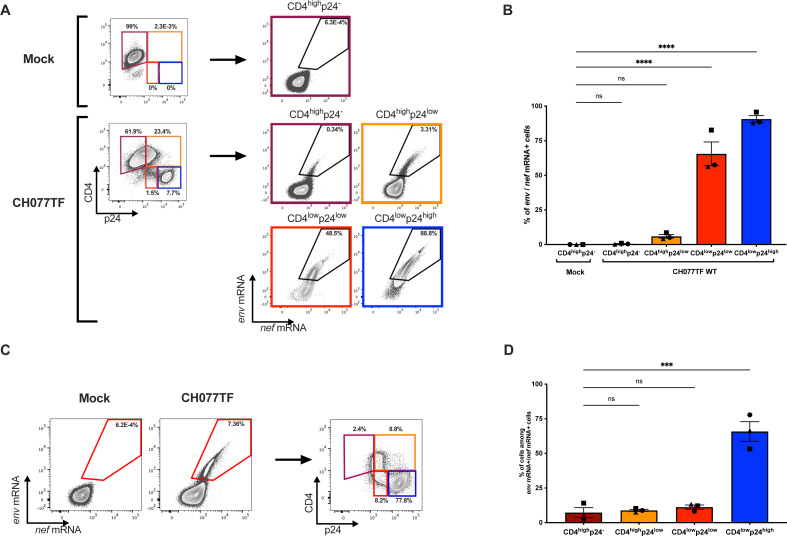
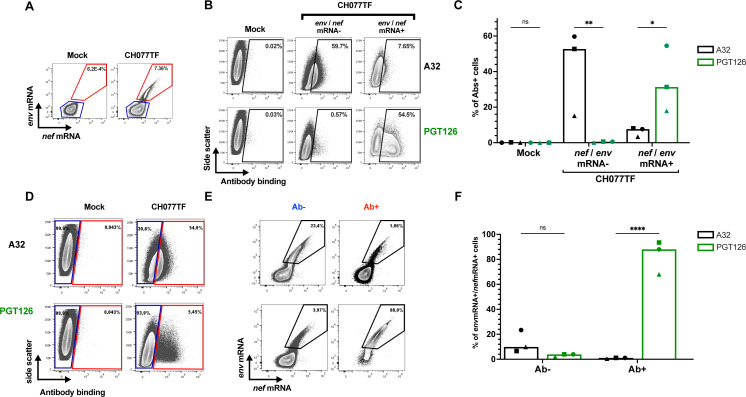
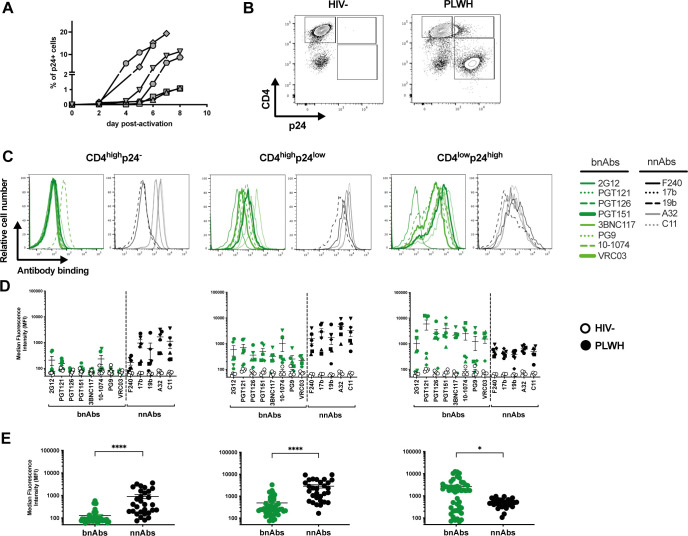
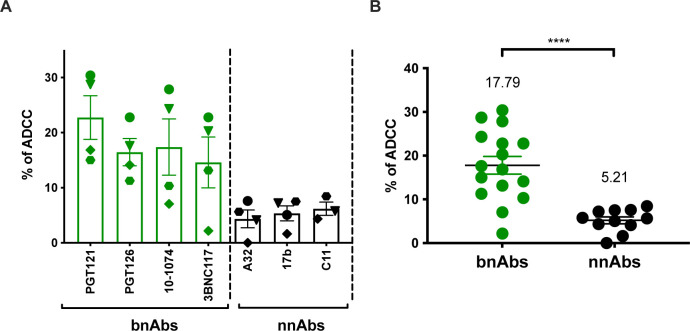
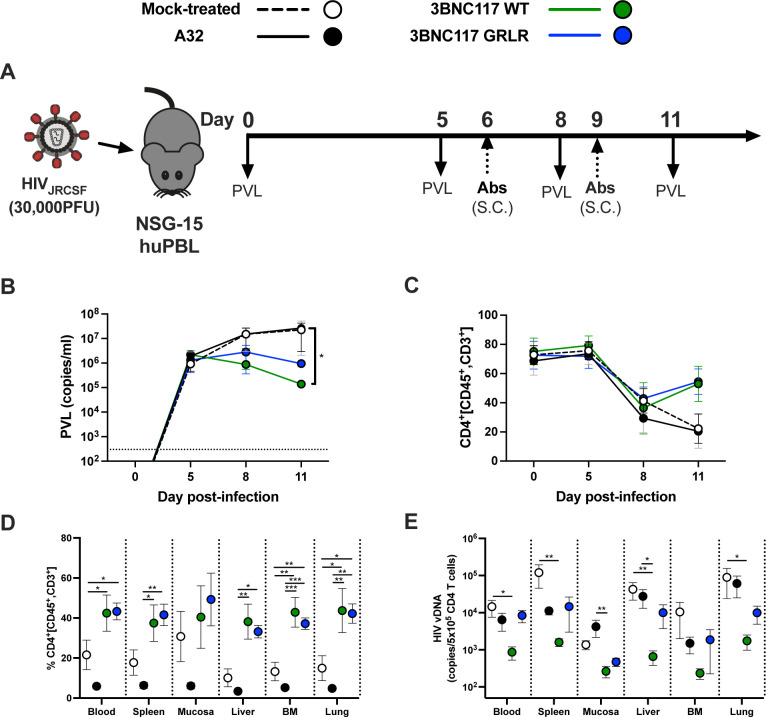
HIV-1 envelope glycoprotein (Env) conformation substantially impacts antibody-dependent cellular cytotoxicity (ADCC). Envs from primary HIV-1 isolates adopt a prefusion "closed" conformation, which is targeted by broadly neutralizing antibodies (bnAbs). CD4 binding drives Env into more "open" conformations, which are recognized by non-neutralizing Abs (nnAbs). To better understand Env-Ab and Env-CD4 interaction in CD4+ T cells infected with HIV-1, we simultaneously measured antibody binding and HIV-1 mRNA expression using multiparametric flow cytometry and RNA flow fluorescent in situ hybridization (FISH) techniques. We observed that env mRNA is almost exclusively expressed by HIV-1 productively infected cells that already downmodulated CD4. This suggests that CD4 downmodulation precedes env mRNA expression. Consequently, productively infected cells express "closed" Envs on their surface, which renders them resistant to nnAbs. Cells recognized by nnAbs were all env mRNA negative, indicating Ab binding through shed gp120 or virions attached to their surface. Consistent with these findings, treatment of HIV-1-infected humanized mice with the ADCC-mediating nnAb A32 failed to lower viral replication or reduce the size of the viral reservoir. These findings confirm the resistance of productively infected CD4+ T cells to nnAbs-mediated ADCC and question the rationale of immunotherapy approaches using this strategy.
Importance: Antibody-dependent cellular cytotoxicity (ADCC) represents an effective immune response for clearing virally infected cells, making ADCC-mediating antibodies promising therapeutic candidates for HIV-1 cure strategies. Broadly neutralizing antibodies (bNAbs) target epitopes present on the native "closed" envelope glycoprotein (Env), while non-neutralizing antibodies (nnAbs) recognize epitopes exposed upon Env-CD4 interaction. Here, we provide evidence that env mRNA is predominantly expressed by productively infected cells that have already downmodulated cell-surface CD4. This indicates that CD4 downmodulation by HIV-1 precedes Env expression, making productively infected cells resistant to ADCC mediated by nnAbs but sensitive to those mediated by bnAbs. These findings offer critical insights for the development of immunotherapy-based strategies aimed at targeting and eliminating productively infected cells in people living with HIV.
Keywords: A32; ADCC; Env; HIV-1; RNA-flow fish; bNAbs; hu-mice; nnAbs.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The combination of three CD4-induced antibodies targeting highly conserved Env regions with a small CD4-mimetic achieves potent ADCC activity.J Virol. 2024 Oct 22;98(10):e0101624. doi: 10.1128/jvi.01016-24. Epub 2024 Sep 9. J Virol. 2024. PMID: 39248460
-
Antibody-Dependent Cellular Cytotoxicity-Competent Antibodies against HIV-1-Infected Cells in Plasma from HIV-Infected Subjects.mBio. 2019 Dec 17;10(6):e02690-19. doi: 10.1128/mBio.02690-19. mBio. 2019. PMID: 31848282 Free PMC article.
-
Interaction with cellular CD4 exposes HIV-1 envelope epitopes targeted by antibody-dependent cell-mediated cytotoxicity.J Virol. 2014 Mar;88(5):2633-44. doi: 10.1128/JVI.03230-13. Epub 2013 Dec 18. J Virol. 2014. PMID: 24352444 Free PMC article.
-
Unlocking HIV-1 Env: implications for antibody attack.AIDS Res Ther. 2017 Sep 12;14(1):42. doi: 10.1186/s12981-017-0168-5. AIDS Res Ther. 2017. PMID: 28893275 Free PMC article. Review.
-
Impact of HIV-1 Envelope Conformation on ADCC Responses.Trends Microbiol. 2018 Apr;26(4):253-265. doi: 10.1016/j.tim.2017.10.007. Epub 2017 Nov 20. Trends Microbiol. 2018. PMID: 29162391 Review.
Cited by
-
Three families of CD4-induced antibodies are associated with the capacity of plasma from people living with HIV to mediate ADCC in presence of CD4-mimetics.medRxiv [Preprint]. 2024 Jun 4:2024.06.02.24308281. doi: 10.1101/2024.06.02.24308281. medRxiv. 2024. Update in: J Virol. 2024 Oct 22;98(10):e0096024. doi: 10.1128/jvi.00960-24 PMID: 38883797 Free PMC article. Updated. Preprint.
-
The combination of three CD4-induced antibodies targeting highly conserved Env regions with a small CD4-mimetic achieves potent ADCC activity.bioRxiv [Preprint]. 2024 Jun 7:2024.06.07.597978. doi: 10.1101/2024.06.07.597978. bioRxiv. 2024. Update in: J Virol. 2024 Oct 22;98(10):e0101624. doi: 10.1128/jvi.01016-24 PMID: 38895270 Free PMC article. Updated. Preprint.
-
Three families of CD4-induced antibodies are associated with the capacity of plasma from people living with HIV to mediate ADCC in the presence of CD4-mimetics.J Virol. 2024 Oct 22;98(10):e0096024. doi: 10.1128/jvi.00960-24. Epub 2024 Sep 4. J Virol. 2024. PMID: 39230306
-
The asymmetric opening of HIV-1 Env by a potent CD4 mimetic enables anti-coreceptor binding site antibodies to mediate ADCC.bioRxiv [Preprint]. 2024 Aug 27:2024.08.27.609961. doi: 10.1101/2024.08.27.609961. bioRxiv. 2024. PMID: 39253431 Free PMC article. Preprint.
-
The combination of three CD4-induced antibodies targeting highly conserved Env regions with a small CD4-mimetic achieves potent ADCC activity.J Virol. 2024 Oct 22;98(10):e0101624. doi: 10.1128/jvi.01016-24. Epub 2024 Sep 9. J Virol. 2024. PMID: 39248460
References
-
- Finzi D, Hermankova M, Pierson T, Carruth LM, Buck C, Chaisson RE, Quinn TC, Chadwick K, Margolick J, Brookmeyer R, Gallant J, Markowitz M, Ho DD, Richman DD, Siliciano RF. 1997. Identification of a reservoir for HIV-1 in patients on highly active antiretroviral therapy. Science 278:1295–1300. doi:10.1126/science.278.5341.1295 - DOI - PubMed
-
- Chomont N, El-Far M, Ancuta P, Trautmann L, Procopio FA, Yassine-Diab B, Boucher G, Boulassel M-R, Ghattas G, Brenchley JM, Schacker TW, Hill BJ, Douek DC, Routy J-P, Haddad EK, Sékaly R-P. 2009. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat Med 15:893–900. doi:10.1038/nm.1972 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- UM1 AI144462/AI/NIAID NIH HHS/United States
- R01 AI148379/AI/NIAID NIH HHS/United States
- R01 AI129769/AI/NIAID NIH HHS/United States
- UM1 AI164570/AI/NIAID NIH HHS/United States
- R01 AI150322/AI/NIAID NIH HHS/United States
- R01 AI145164/AI/NIAID NIH HHS/United States
- P01 GM056550/GM/NIGMS NIH HHS/United States
- R01 AI162646/AI/NIAID NIH HHS/United States
- R01 AI186809/AI/NIAID NIH HHS/United States
- UM1 AI164562/AI/NIAID NIH HHS/United States
- UM1 AI144371/AI/NIAID NIH HHS/United States
- P01 AI150471/AI/NIAID NIH HHS/United States
- R01 AI176531/AI/NIAID NIH HHS/United States
- UM1 AI164559/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials
