

This is a preprint.
Structural Analyses of a GABARAP~ATG3 Conjugate Uncover a Novel Non-covalent Ubl-E2 Backside Interaction
- PMID: 39185234
- PMCID: PMC11343110
- DOI: 10.1101/2024.08.14.607425
Structural Analyses of a GABARAP~ATG3 Conjugate Uncover a Novel Non-covalent Ubl-E2 Backside Interaction
Abstract
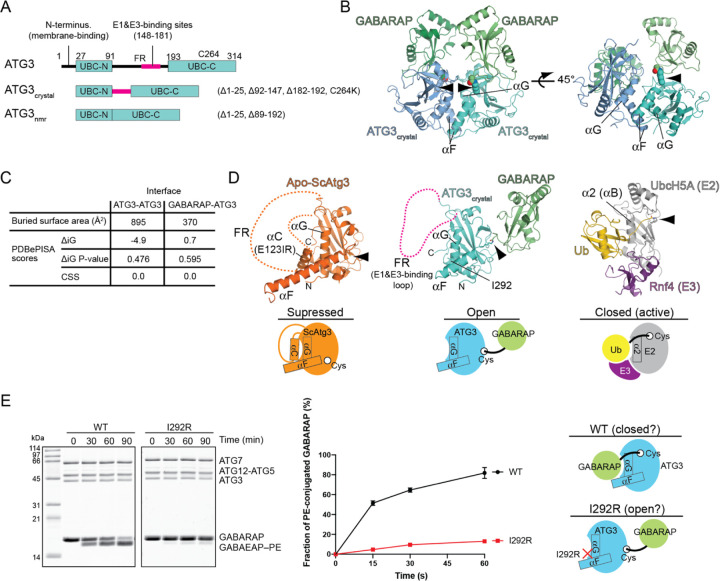
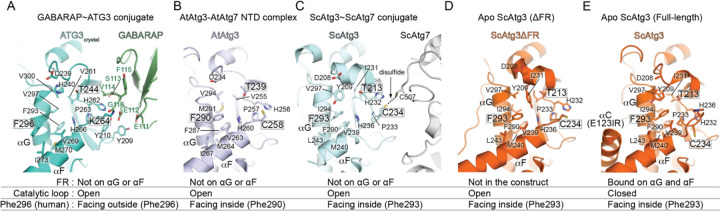
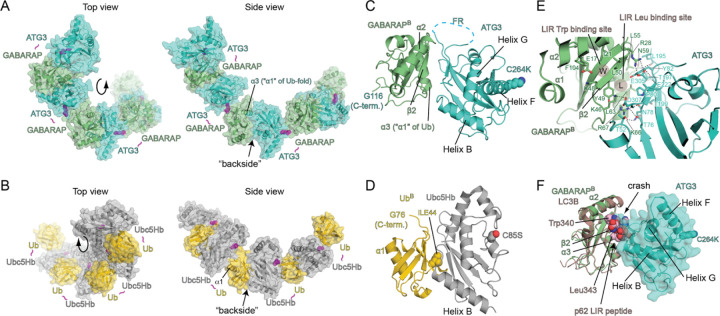
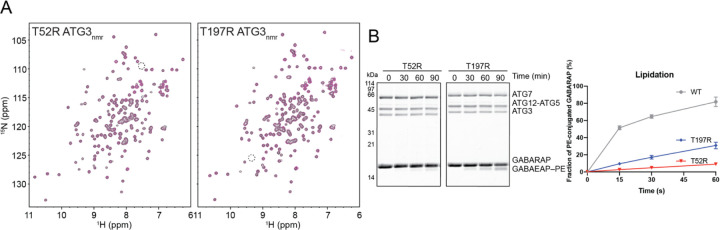
Members of the ATG8 family of ubiquitin-like proteins (Ubls) are conjugated to phosphatidylethanolamine (PE) in the autophagosomal membrane, where they recruit degradation substrates and facilitate membrane biogenesis. Despite this well-characterized function, the mechanisms underlying the lipidation process, including the action of the E2 enzyme ATG3, remain incompletely understood. Here, we report the crystal structure of human ATG3 conjugated to the mammalian ATG8 protein GABARAP via an isopeptide bond, mimicking the Ubl~E2 thioester intermediate. In this structure, the GABARAP~ATG3 conjugate adopts an open configuration with minimal contacts between the two proteins. Notably, the crystal lattice reveals non-covalent contacts between GABARAP and the backside of ATG3's E2 catalytic center, resulting in the formation of a helical filament of the GABARAP~ATG3 conjugate. While similar filament formations have been observed with canonical Ub~E2 conjugates, the E2 backside-binding interface of GABARAP is distinct from those of Ub/Ubl proteins and overlaps with the binding site for LC3 interacting region (LIR) peptides. NMR analysis confirms the presence of this non-covalent interaction in solution, and mutagenesis experiments demonstrate the involvement of the E2 backside in PE conjugation. These findings highlight the critical role of the E2 backside in the lipidation process and suggest evolutionary adaptations in the unique E2 enzyme ATG3.
Figures


Similar articles
-
Production of Human ATG Proteins for Lipidation Assays.Methods Enzymol. 2017;587:97-113. doi: 10.1016/bs.mie.2016.09.055. Epub 2016 Nov 11. Methods Enzymol. 2017. PMID: 28253979 Free PMC article.
-
A switch element in the autophagy E2 Atg3 mediates allosteric regulation across the lipidation cascade.Nat Commun. 2019 Aug 9;10(1):3600. doi: 10.1038/s41467-019-11435-y. Nat Commun. 2019. PMID: 31399562 Free PMC article.
-
Allosteric regulation through a switch element in the autophagy E2, Atg3.Autophagy. 2020 Jan;16(1):183-184. doi: 10.1080/15548627.2019.1688550. Epub 2019 Nov 9. Autophagy. 2020. PMID: 31690182 Free PMC article. Review.
-
Human ATG4 autophagy proteases counteract attachment of ubiquitin-like LC3/GABARAP proteins to other cellular proteins.J Biol Chem. 2019 Aug 23;294(34):12610-12621. doi: 10.1074/jbc.AC119.009977. Epub 2019 Jul 17. J Biol Chem. 2019. PMID: 31315929 Free PMC article.
-
In vitro assays of lipidation of Mammalian Atg8 homologs.Curr Protoc Cell Biol. 2014 Sep 2;64:11.20.1-13. doi: 10.1002/0471143030.cb1120s64. Curr Protoc Cell Biol. 2014. PMID: 25181299 Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources