

Released bacterial ATP shapes local and systemic inflammation during abdominal sepsis
- PMID: 39163101
- PMCID: PMC11335348
- DOI: 10.7554/eLife.96678
Released bacterial ATP shapes local and systemic inflammation during abdominal sepsis
Abstract
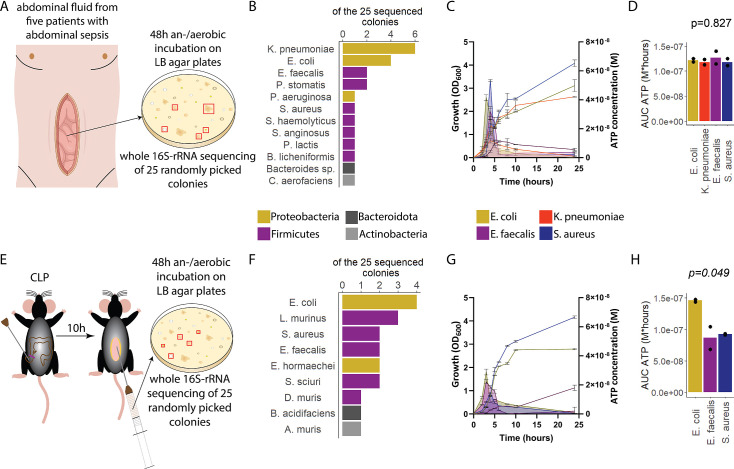
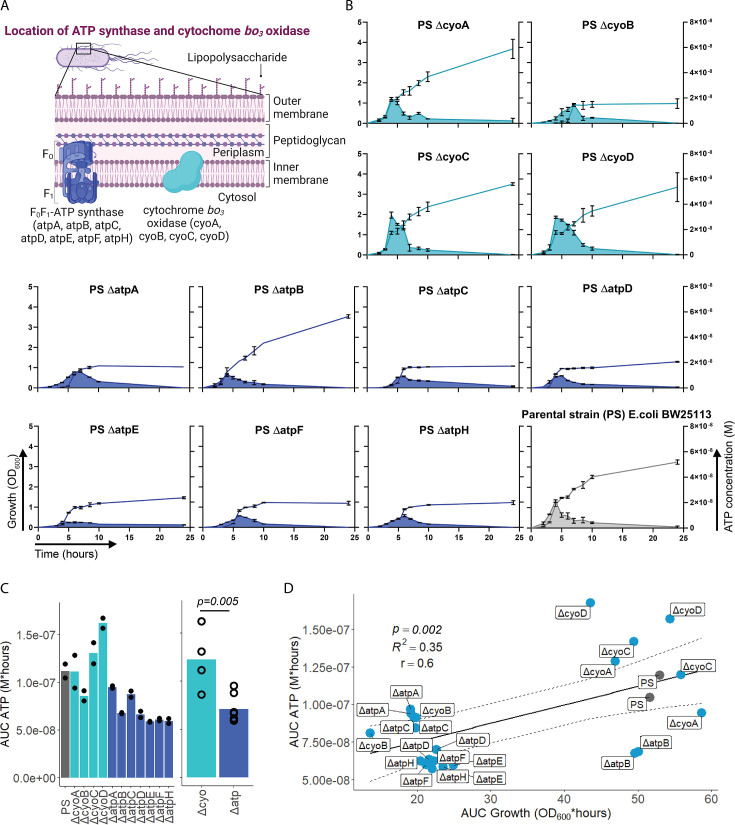
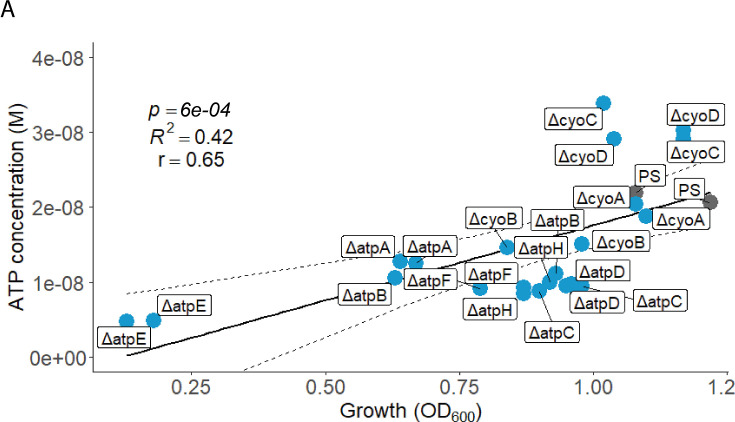
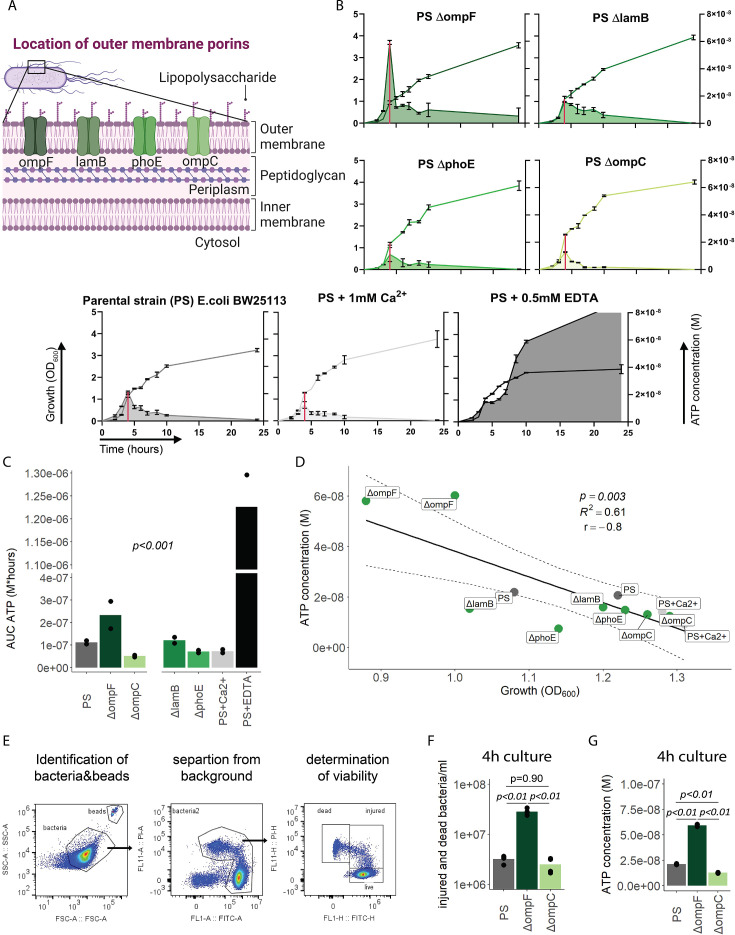
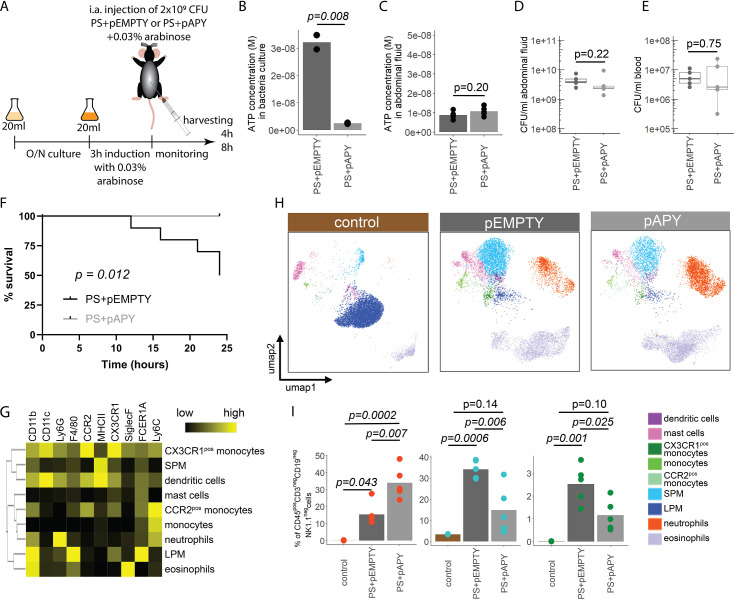
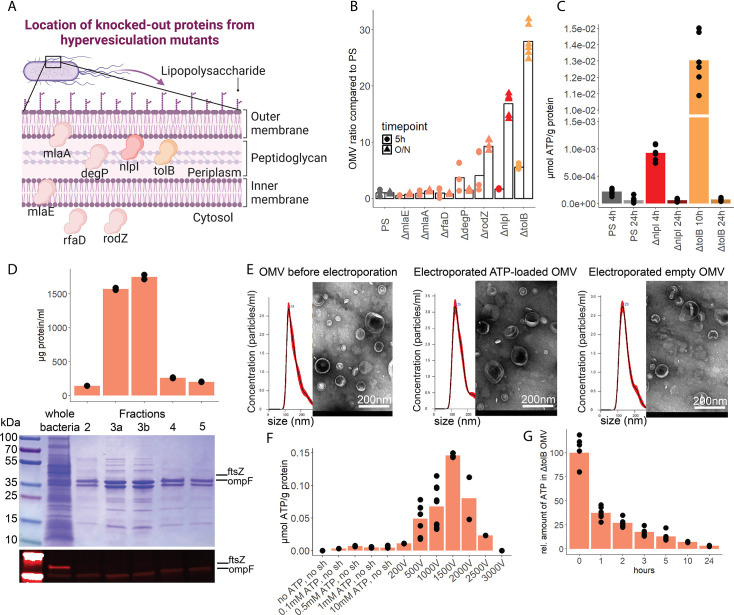
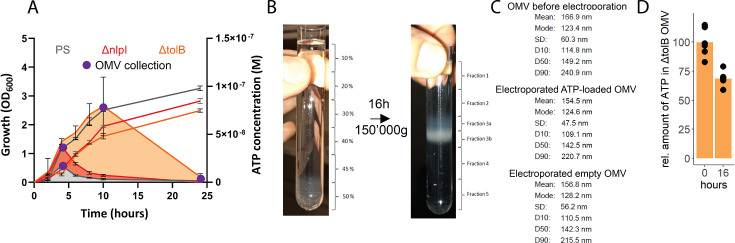
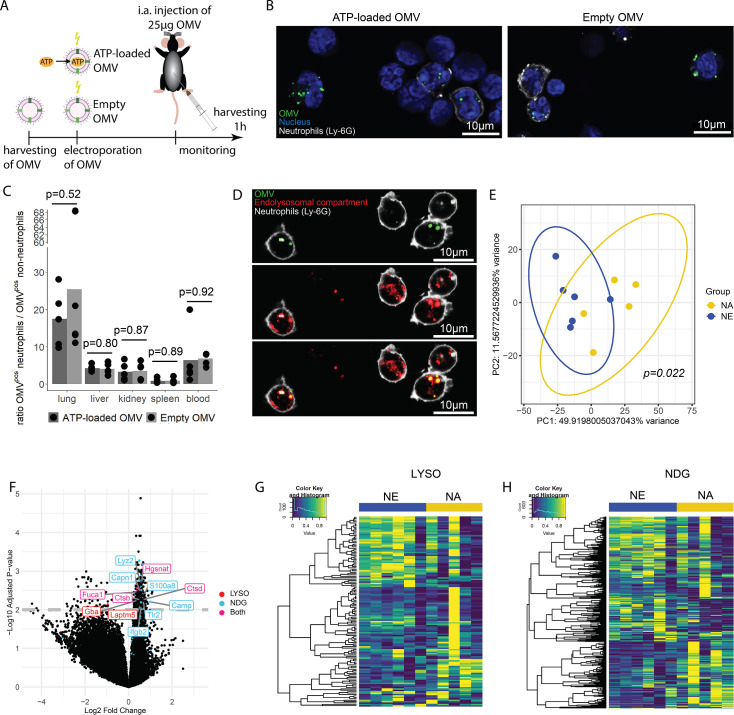
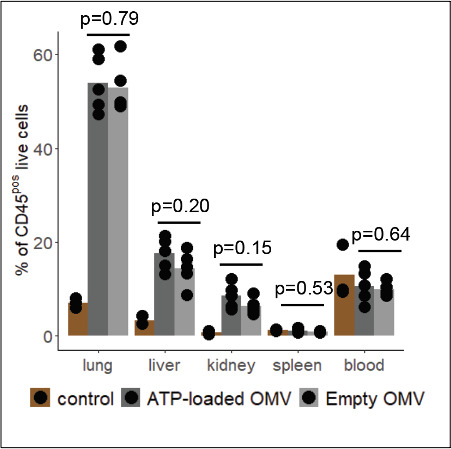
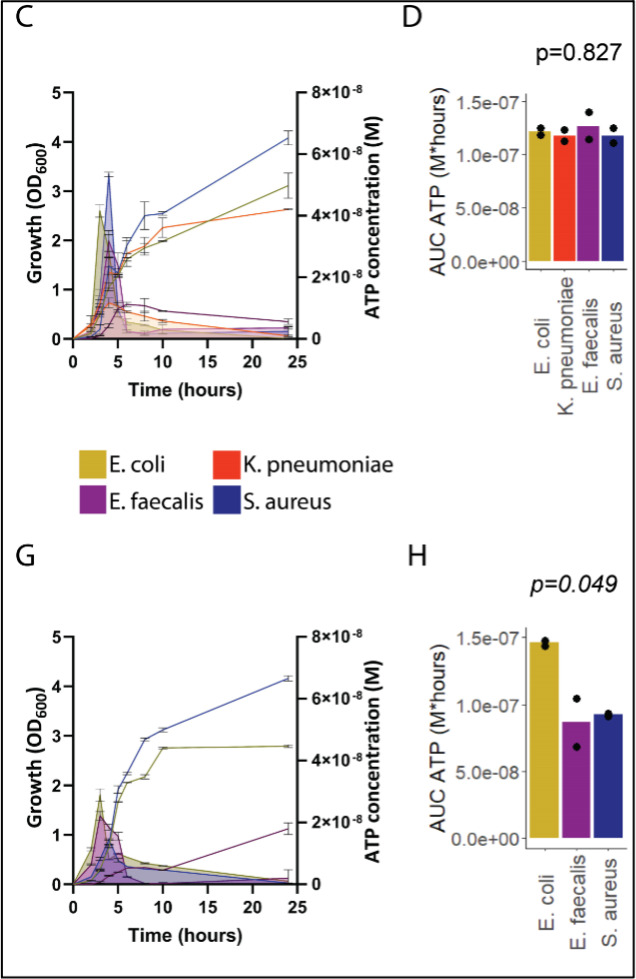
Sepsis causes millions of deaths per year worldwide and is a current global health priority declared by the WHO. Sepsis-related deaths are a result of dysregulated inflammatory immune responses indicating the need to develop strategies to target inflammation. An important mediator of inflammation is extracellular adenosine triphosphate (ATP) that is released by inflamed host cells and tissues, and also by bacteria in a strain-specific and growth-dependent manner. Here, we investigated the mechanisms by which bacteria release ATP. Using genetic mutant strains of Escherichia coli (E. coli), we demonstrate that ATP release is dependent on ATP synthase within the inner bacterial membrane. In addition, impaired integrity of the outer bacterial membrane notably contributes to ATP release and is associated with bacterial death. In a mouse model of abdominal sepsis, local effects of bacterial ATP were analyzed using a transformed E. coli bearing an arabinose-inducible periplasmic apyrase hydrolyzing ATP to be released. Abrogating bacterial ATP release shows that bacterial ATP suppresses local immune responses, resulting in reduced neutrophil counts and impaired survival. In addition, bacterial ATP has systemic effects via its transport in outer membrane vesicles (OMV). ATP-loaded OMV are quickly distributed throughout the body and upregulated expression of genes activating degranulation in neutrophils, potentially contributing to the exacerbation of sepsis severity. This study reveals mechanisms of bacterial ATP release and its local and systemic roles in sepsis pathogenesis.
Keywords: ATP; E. coli; OMV; bacterial ATP; immunology; infectious disease; inflammation; microbiology; mouse; sepsis.
Plain language summary
Sepsis is a severe condition often caused by the body’s immune system overreacting to bacterial infections. This can lead to excessive inflammation which damages organs and requires urgent medical care. With sepsis claiming millions of lives each year, new and improved ways to treat this condition are urgently needed. One potential strategy for treating sepsis is to target the underlying mechanisms controlling inflammation. Inflamed and dying cells release molecules called ATP (the energy carrier of all living cells), which strongly influence the immune system, including during sepsis. In the early stages of an infection, ATP acts as a danger signal warning the body that something is wrong. However, over time, it can worsen infections by disturbing the immune response. Similar to human cells, bacteria release their own ATP, which can have different impacts depending on the type of bacteria and where they are located in the body. However, it is not well understood how bacterial ATP influences severe infections like sepsis. To investigate this question, Spari et al analysed how ATP is released from Escherichia coli, a type of bacteria that causes severe infections. This revealed that the bacteria secrete ATP directly in to their environment and via small membrane-bound structures called vesicles. Spari et al. then probed a mouse model of abdominal sepsis which had been infected with E. coli that release either normal or low levels of ATP. They found that the ATP released from E. coli impaired the mice’s survival and lowered the number of neutrophils (immune cells which are important for defending against bacteria) at the site of the infection. The ATP secreted via vesicles also altered the role of neutrophils but in more distant regions, and it is possible that these changes may be contributing to the severity of sepsis. These findings provide a better understanding of how ATP released from bacteria impacts the immune system during sepsis. While further investigation is needed, these findings may offer new therapeutic targets for treating sepsis.
© 2024, Spari et al.
Conflict of interest statement
DS, AS, DS, SM, KK, NE, LS, DS, GB No competing interests declared
Figures







Update of
- doi: 10.1101/2024.03.07.583973
- doi: 10.7554/eLife.96678.1
- doi: 10.7554/eLife.96678.2
Similar articles
-
CD14 release induced by P2X7 receptor restricts inflammation and increases survival during sepsis.Elife. 2020 Nov 2;9:e60849. doi: 10.7554/eLife.60849. Elife. 2020. PMID: 33135636 Free PMC article.
-
Frontline Science: Escherichia coli use LPS as decoy to impair neutrophil chemotaxis and defeat antimicrobial host defense.J Leukoc Biol. 2019 Dec;106(6):1211-1219. doi: 10.1002/JLB.4HI0319-109R. Epub 2019 Aug 8. J Leukoc Biol. 2019. PMID: 31392789 Free PMC article.
-
Toll-like receptor-triggered calcium mobilization protects mice against bacterial infection through extracellular ATP release.Infect Immun. 2014 Dec;82(12):5076-85. doi: 10.1128/IAI.02546-14. Epub 2014 Sep 22. Infect Immun. 2014. PMID: 25245808 Free PMC article.
-
Secretion and Delivery of Intestinal Pathogenic Escherichia coli Virulence Factors via Outer Membrane Vesicles.Front Cell Infect Microbiol. 2020 Mar 6;10:91. doi: 10.3389/fcimb.2020.00091. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32211344 Free PMC article. Review.
-
The bacteria and the host: a story of purinergic signaling in urinary tract infections.Am J Physiol Cell Physiol. 2021 Jul 1;321(1):C134-C146. doi: 10.1152/ajpcell.00054.2021. Epub 2021 May 12. Am J Physiol Cell Physiol. 2021. PMID: 33979212 Review.
Cited by
-
Horizontal gene transfer and beyond: the delivery of biological matter by bacterial membrane vesicles to host and bacterial cells.Curr Opin Microbiol. 2024 Oct;81:102525. doi: 10.1016/j.mib.2024.102525. Epub 2024 Aug 26. Curr Opin Microbiol. 2024. PMID: 39190937 Review.
References
-
- Arbizu PM. pairwiseAdonis. cb190f7GitHub. 2023 https://github.com/pmartinezarbizu/pairwiseAdonis
MeSH terms
Substances
Associated data
- Actions
