

Immunological characteristics of a recombinant alphaherpesvirus with an envelope-embedded Cap protein of circovirus
- PMID: 39081314
- PMCID: PMC11286414
- DOI: 10.3389/fimmu.2024.1438371
Immunological characteristics of a recombinant alphaherpesvirus with an envelope-embedded Cap protein of circovirus
Erratum in
-
Corrigendum: Immunological characteristics of a recombinant alphaherpesvirus with an envelope-embedded Cap protein of circovirus.Front Immunol. 2024 Nov 18;15:1515309. doi: 10.3389/fimmu.2024.1515309. eCollection 2024. Front Immunol. 2024. PMID: 39624086 Free PMC article.
Abstract
Introduction: Variant pseudorabies virus (PRV) is a newly emerged zoonotic pathogen that can cause human blindness. PRV can take advantage of its large genome and multiple non-essential genes to construct recombinant attenuated vaccines carrying foreign genes. However, a major problem is that the foreign genes in recombinant PRV are only integrated into the genome for independent expression, rather than assembled on the surface of virion.
Methods: We reported a recombinant PRV with deleted gE/TK genes and an inserted porcine circovirus virus 2 (PCV2) Cap gene into the extracellular domain of the PRV gE gene using the Cre-loxP recombinant system combined with the CRISPR-Cas9 gene editing system. This recombinant PRV (PRV-Cap), with the envelope-embedded Cap protein, exhibits a similar replication ability to its parental virus.
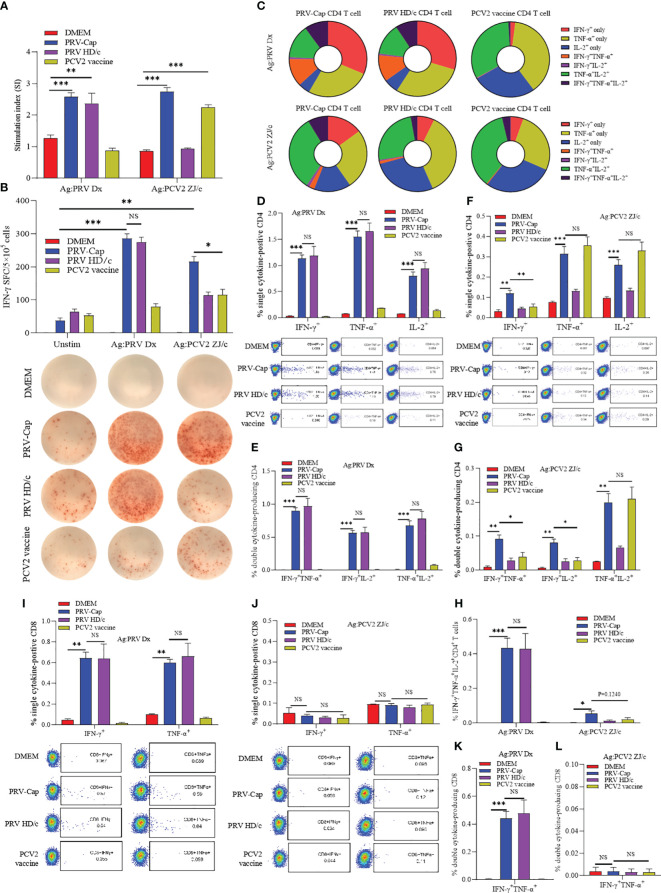
Results: An immunogenicity assay revealed that PRV-Cap immunized mice have 100% resistance to lethal PRV and PCV2 attacks. Neutralization antibody and ELISPOT detections indicated that PRV-Cap can enhance neutralizing antibodies to PRV and produce IFN-γ secreting T cells specific for both PRV and PCV2. Immunological mechanistic investigation revealed that initial immunization with PRV-Cap stimulates significantly early activation and expansion of CD69+ T cells, promoting the activation of CD4 Tfh cell dependent germinal B cells and producing effectively specific effector memory T and B cells. Booster immunization with PRV-Cap recalled the activation of PRV-specific IFN-γ+IL-2+CD4+ T cells and IFN-γ+TNF-α+CD8+ T cells, as well as PCV2-specific IFN-γ+TNF-α+CD8+ T cells.
Conclusion: Collectively, our data suggested an immunological mechanism in that the recombinant PRV with envelope-assembled PCV2 Cap protein can serve as an excellent vaccine candidate for combined immunity against PRV and PCV2, and provided a cost-effective method for the production of PRV- PCV2 vaccine.
Keywords: IFN-γ; chimeric pseudorabies virus; circovirus; immunity; memory responses.
Copyright © 2024 Lu, Li, Chen, Li, Ma, Peng, Yan, Dong, Jin, Pan, Shang, Gu and Zhou.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
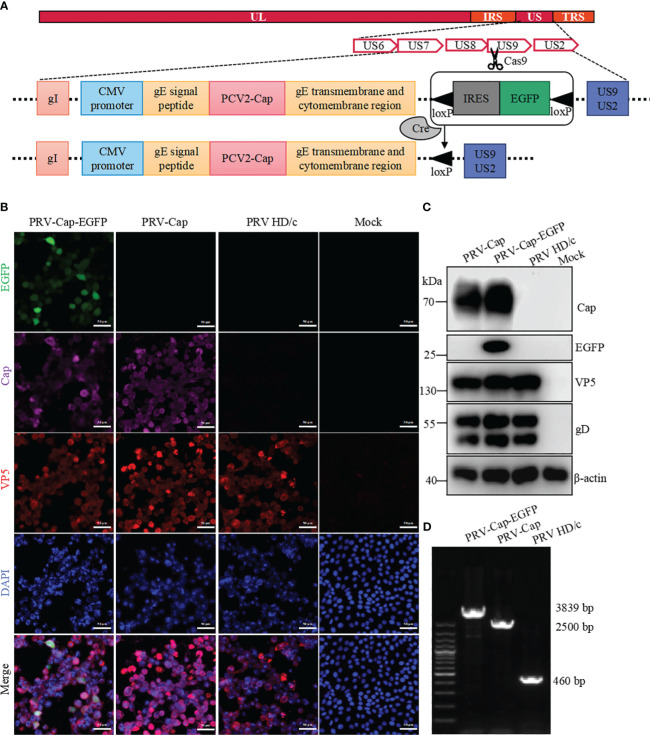
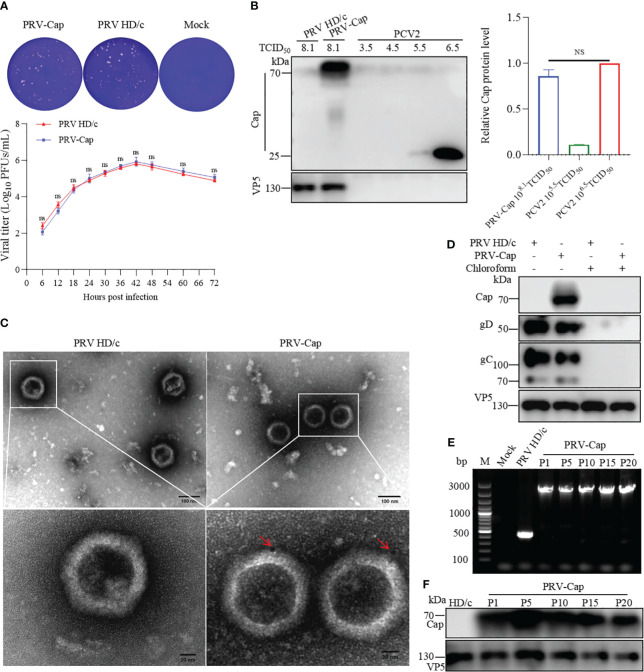
Similar articles
-
Depressing time: Waiting, melancholia, and the psychoanalytic practice of care.In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. In: Kirtsoglou E, Simpson B, editors. The Time of Anthropology: Studies of Contemporary Chronopolitics. Abingdon: Routledge; 2020. Chapter 5. PMID: 36137063 Free Books & Documents. Review.
-
Optimizing encephalomyocarditis virus VP1 protein assembly on pseudorabies virus envelope via US9 protein anchoring.Virulence. 2025 Dec;16(1):2445235. doi: 10.1080/21505594.2024.2445235. Epub 2024 Dec 27. Virulence. 2025. PMID: 39727074 Free PMC article.
-
Different immunogens and prime-boost vaccination strategies affect the efficacy of recombinant candidate vaccines against pathogenic orthopoxviruses.Virol J. 2024 Nov 7;21(1):282. doi: 10.1186/s12985-024-02534-4. Virol J. 2024. PMID: 39511612 Free PMC article.
-
A broadly neutralizing human monoclonal antibody generated from transgenic mice immunized with HCMV particles limits virus infection and proliferation.J Virol. 2024 Jul 23;98(7):e0021324. doi: 10.1128/jvi.00213-24. Epub 2024 Jun 4. J Virol. 2024. PMID: 38832789 Free PMC article.
-
Continuing education meetings and workshops: effects on professional practice and healthcare outcomes.Cochrane Database Syst Rev. 2021 Sep 15;9(9):CD003030. doi: 10.1002/14651858.CD003030.pub3. Cochrane Database Syst Rev. 2021. PMID: 34523128 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous