

ESCRT-III: a versatile membrane remodeling machinery and its implications in cellular processes and diseases
- PMID: 39070887
- PMCID: PMC11275535
- DOI: 10.1080/19768354.2024.2380294
ESCRT-III: a versatile membrane remodeling machinery and its implications in cellular processes and diseases
Abstract
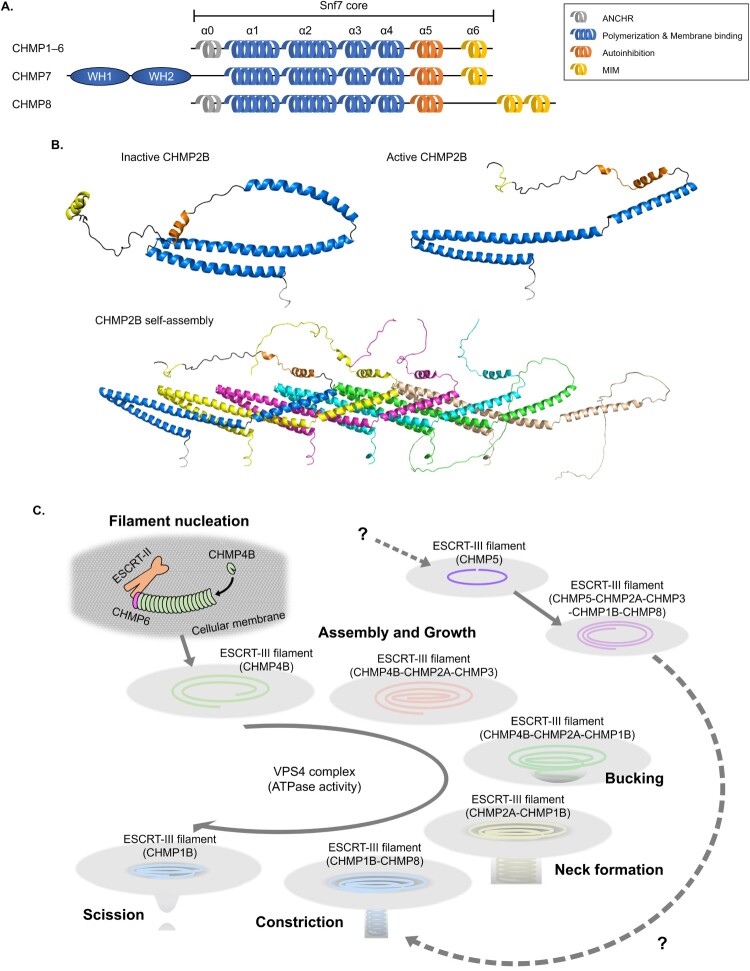
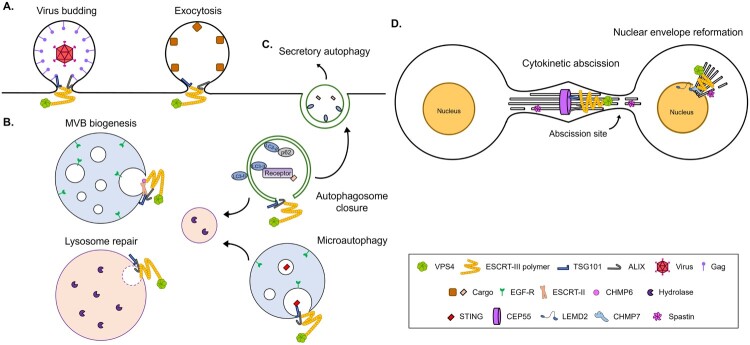
The endosomal sorting complexes required for transport (ESCRT) machinery is an evolutionarily conserved cytosolic protein complex that plays a crucial role in membrane remodeling and scission events across eukaryotes. Initially discovered for its function in multivesicular body (MVB) formation, the ESCRT complex has since been implicated in a wide range of membrane-associated processes, including endocytosis, exocytosis, cytokinesis, and autophagy. Recent advances have elucidated the ESCRT assembly pathway and highlighted the distinct functions of the various ESCRT complexes and their associated partners. Among the ESCRT complexes, ESCRT-III stands out as a critical player in membrane remodeling, with its subunits assembled into higher-order multimers capable of bending and severing membranes. This review focuses on the ESCRT-III complex, exploring its diverse functions in cellular processes beyond MVB biogenesis. We delve into the molecular mechanisms underlying ESCRT-III-mediated membrane remodeling and highlight its emerging roles in processes such as viral budding, autophagosome closure, and cytokinetic abscission. We also discuss the implications of ESCRT-III dysregulation in neurodegenerative diseases. The versatile membrane remodeling capabilities of ESCRT-III across diverse cellular processes underscore its importance in maintaining proper cellular function. Furthermore, we highlight the promising potential of ESCRT-III as a therapeutic target for neurodegenerative diseases, offering insights into the treatments of the diseases and the technical applications in related research fields.
Keywords: Autophagy; ESCRT-III; MVB; membrane remodeling; neurodegenerative disease.
© 2024 The Author(s). Published by Informa UK Limited, trading as Taylor & Francis Group.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
Similar articles
-
Cellular Functions and Molecular Mechanisms of the ESCRT Membrane-Scission Machinery.Trends Biochem Sci. 2017 Jan;42(1):42-56. doi: 10.1016/j.tibs.2016.08.016. Epub 2016 Sep 23. Trends Biochem Sci. 2017. PMID: 27669649 Review.
-
Molecular mechanisms of the membrane sculpting ESCRT pathway.Cold Spring Harb Perspect Biol. 2013 Sep 1;5(9):a016766. doi: 10.1101/cshperspect.a016766. Cold Spring Harb Perspect Biol. 2013. PMID: 24003212 Free PMC article. Review.
-
Plant ESCRT Complexes: Moving Beyond Endosomal Sorting.Trends Plant Sci. 2017 Nov;22(11):986-998. doi: 10.1016/j.tplants.2017.08.003. Epub 2017 Aug 31. Trends Plant Sci. 2017. PMID: 28867368 Review.
-
The ESCRT machinery: from the plasma membrane to endosomes and back again.Crit Rev Biochem Mol Biol. 2014 May-Jun;49(3):242-61. doi: 10.3109/10409238.2014.881777. Epub 2014 Jan 24. Crit Rev Biochem Mol Biol. 2014. PMID: 24456136 Free PMC article. Review.
-
A Septin Double Ring Controls the Spatiotemporal Organization of the ESCRT Machinery in Cytokinetic Abscission.Curr Biol. 2019 Jul 8;29(13):2174-2182.e7. doi: 10.1016/j.cub.2019.05.050. Epub 2019 Jun 13. Curr Biol. 2019. PMID: 31204162 Free PMC article.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources